

Inhibition of apoplastic calmodulin impairs calcium homeostasis and cell wall modeling during Cedrus deodara pollen tube growth
- PMID: 23405148
- PMCID: PMC3566176
- DOI: 10.1371/journal.pone.0055411
Inhibition of apoplastic calmodulin impairs calcium homeostasis and cell wall modeling during Cedrus deodara pollen tube growth
Abstract
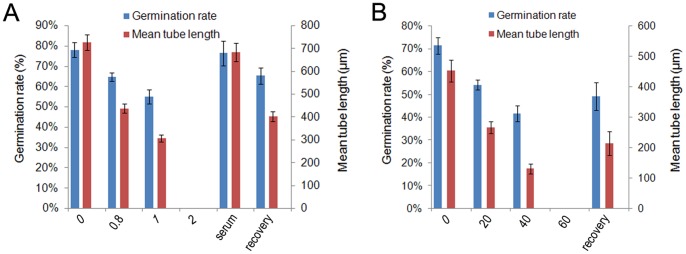
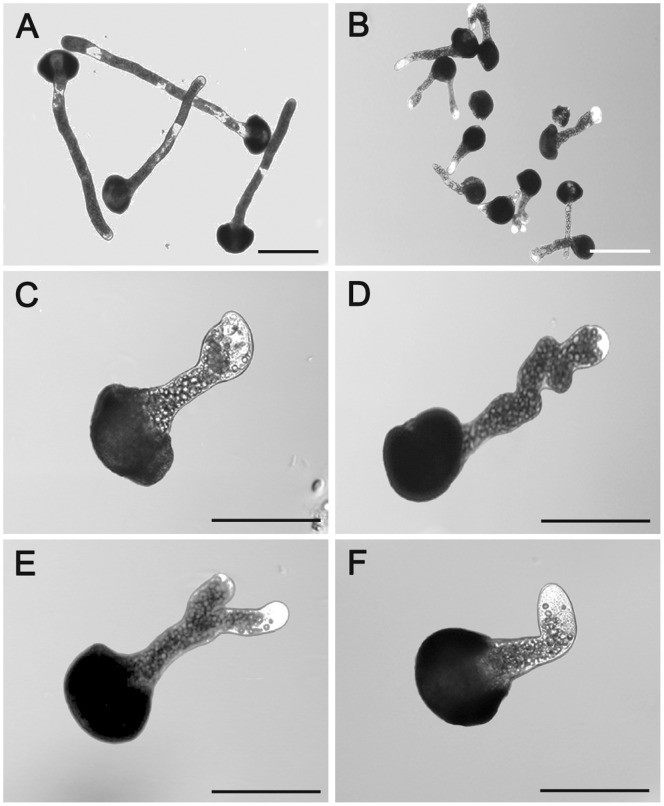
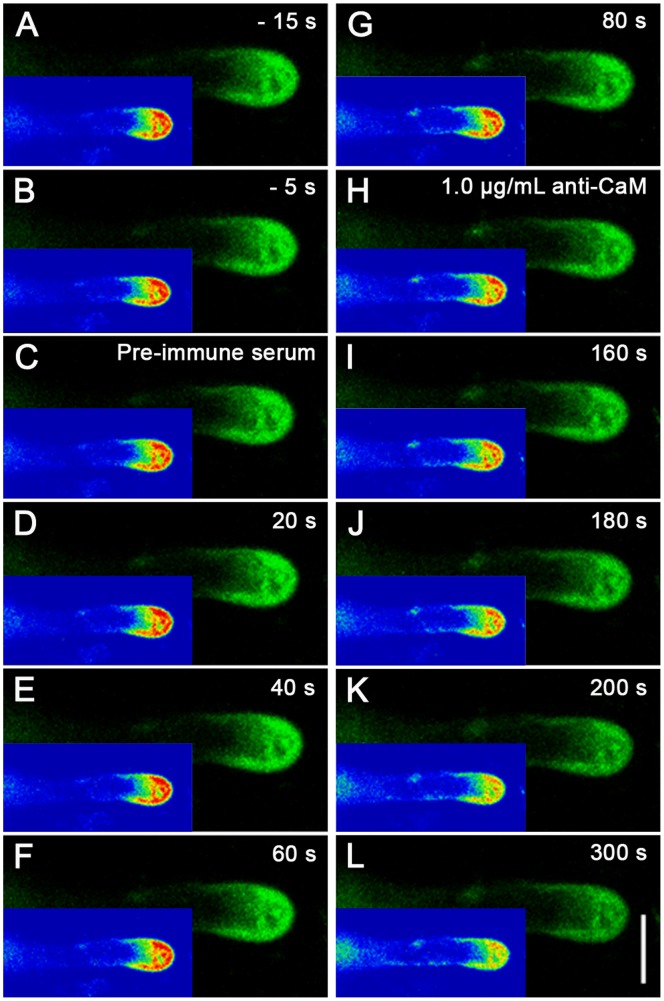
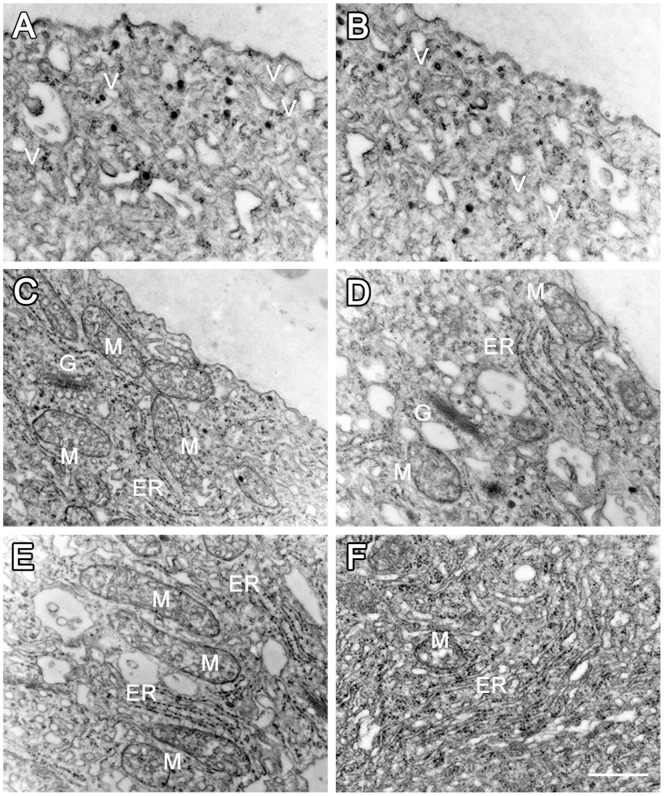
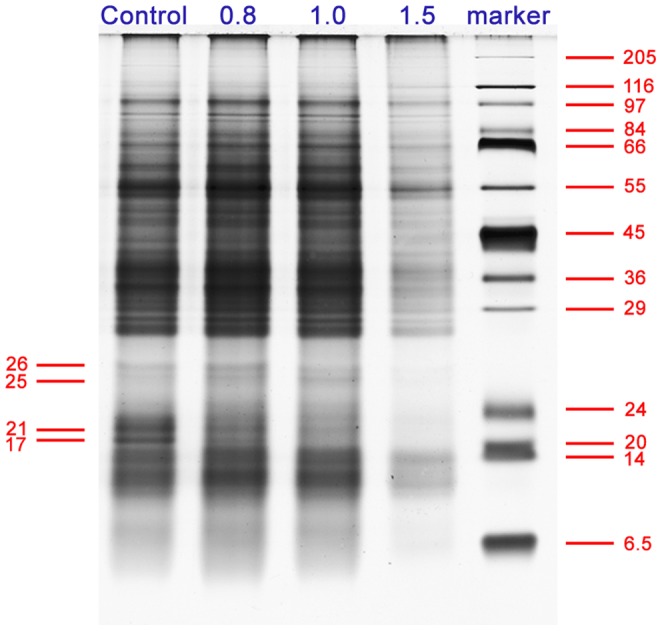
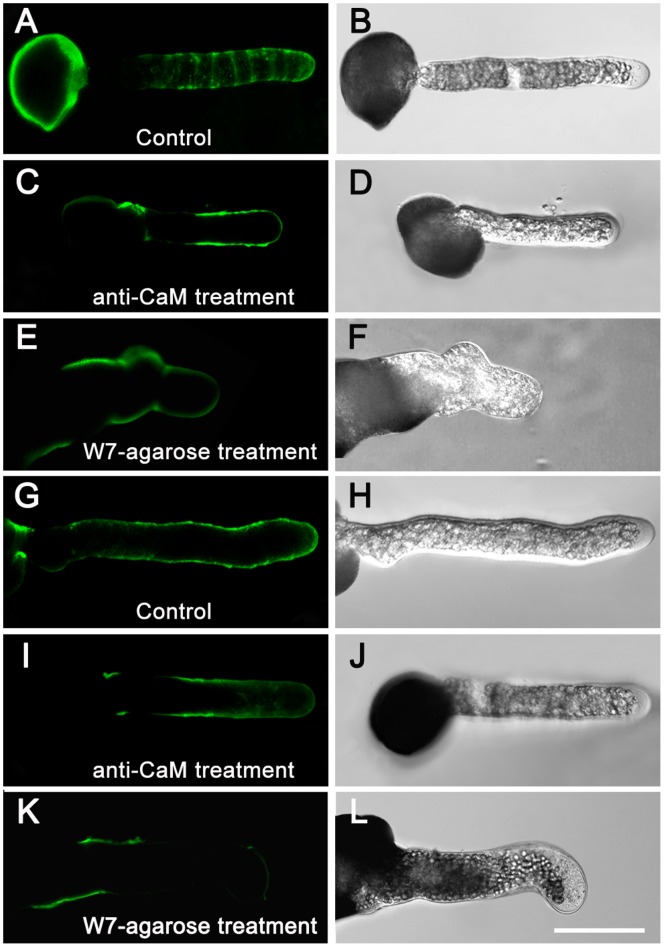
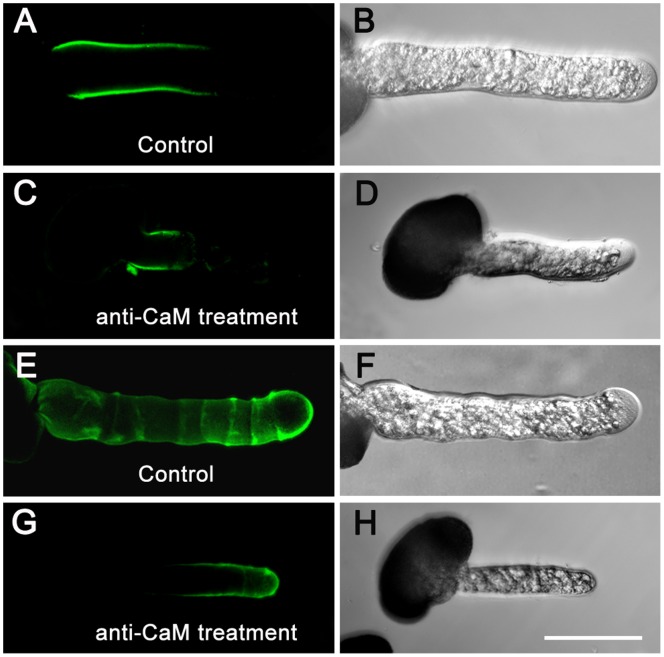
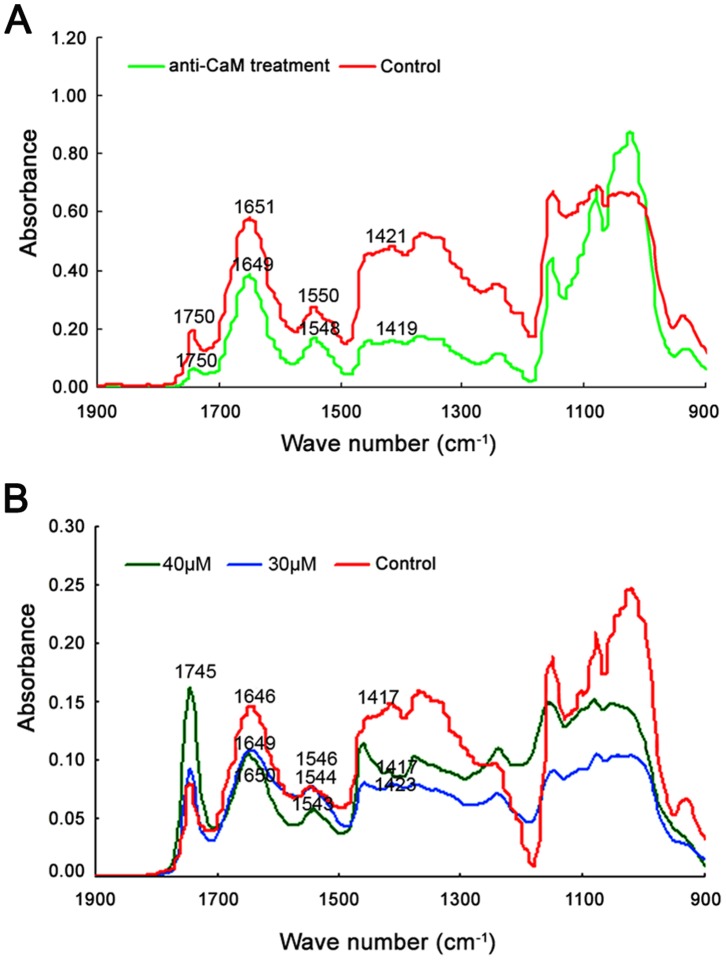
Calmodulin (CaM) is one of the most well-studied Ca(2+) transducers in eukaryotic cells. It is known to regulate the activity of numerous proteins with diverse cellular functions; however, the functions of apoplastic CaM in plant cells are still poorly understood. By combining pharmacological analysis and microscopic techniques, we investigated the involvement of apoplastic CaM in pollen tube growth of Cedrus deodara (Roxb.) Loud. It was found that the tip-focused calcium gradient was rapidly disturbed as one of the early events after application of pharmacological agents, while the cytoplasmic organization was not significantly affected. The deposition and distribution of acidic pectins and esterified pectins were also dramatically changed, further perturbing the normal modeling of the cell wall. Several protein candidates from different functional categories may be involved in the responses to inhibition of apoplastic CaM. These results revealed that apoplastic CaM functions to maintain the tip-focused calcium gradient and to modulate the distribution/transformation of pectins during pollen tube growth.
Conflict of interest statement
Figures
Similar articles
-
Apoplastic calmodulin promotes self-incompatibility pollen tube growth by enhancing calcium influx and reactive oxygen species concentration in Pyrus pyrifolia.Plant Cell Rep. 2014 Feb;33(2):255-63. doi: 10.1007/s00299-013-1526-y. Epub 2013 Oct 22. Plant Cell Rep. 2014. PMID: 24145911
-
Calcium dynamics in tomato pollen tubes using the Yellow Cameleon 3.6 sensor.Plant Reprod. 2018 Jun;31(2):159-169. doi: 10.1007/s00497-017-0317-y. Epub 2017 Dec 13. Plant Reprod. 2018. PMID: 29236154
-
The block of intracellular calcium release affects the pollen tube development of Picea wilsonii by changing the deposition of cell wall components.Protoplasma. 2008;233(1-2):39-49. doi: 10.1007/s00709-008-0310-2. Epub 2008 Aug 26. Protoplasma. 2008. PMID: 18726547
-
Calcium - a central regulator of pollen germination and tube growth.Biochim Biophys Acta. 2013 Jul;1833(7):1573-81. doi: 10.1016/j.bbamcr.2012.10.009. Epub 2012 Oct 13. Biochim Biophys Acta. 2013. PMID: 23072967 Review.
-
The role of pectin in plant morphogenesis.Biosystems. 2012 Sep;109(3):397-402. doi: 10.1016/j.biosystems.2012.04.006. Epub 2012 Apr 24. Biosystems. 2012. PMID: 22554809 Review.
Cited by
-
Putrescine modifies the pollen tube growth of tea (Camellia sinensis) by affecting actin organization and cell wall structure.Protoplasma. 2020 Jan;257(1):89-101. doi: 10.1007/s00709-019-01422-x. Epub 2019 Jul 24. Protoplasma. 2020. PMID: 31342152
-
The cotton MYB108 forms a positive feedback regulation loop with CML11 and participates in the defense response against Verticillium dahliae infection.J Exp Bot. 2016 Mar;67(6):1935-50. doi: 10.1093/jxb/erw016. Epub 2016 Feb 11. J Exp Bot. 2016. PMID: 26873979 Free PMC article.
-
Fertilization in flowering plants: an odyssey of sperm cell delivery.Plant Mol Biol. 2020 May;103(1-2):9-32. doi: 10.1007/s11103-020-00987-z. Epub 2020 Mar 2. Plant Mol Biol. 2020. PMID: 32124177 Review.
-
Apoplastic calmodulin promotes self-incompatibility pollen tube growth by enhancing calcium influx and reactive oxygen species concentration in Pyrus pyrifolia.Plant Cell Rep. 2014 Feb;33(2):255-63. doi: 10.1007/s00299-013-1526-y. Epub 2013 Oct 22. Plant Cell Rep. 2014. PMID: 24145911
References
-
- Feijó JA, Costa SS, Prado AM, Becker JD, Certal AC (2004) Signaling by tips. Curr Opin Plant Biol 7: 589–598. - PubMed
-
- Krichevsky A, Kozlovsky SV, Tian GW, Chen MH, Zaltsman A, et al. (2007) How pollen tube grow. Dev Biol 303: 405–420. - PubMed
-
- Boynton AL, Whitfield JF, MacManus JP (1980) Calmodulin stimulating DNA synthesis by rat live cells. Biochem Bioph Res Co 95: 745–749. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
