

Absence of immune responses with xenogeneic collagen and elastin
- PMID: 23406399
- PMCID: PMC3665304
- DOI: 10.1089/ten.TEA.2012.0394
Absence of immune responses with xenogeneic collagen and elastin
Abstract
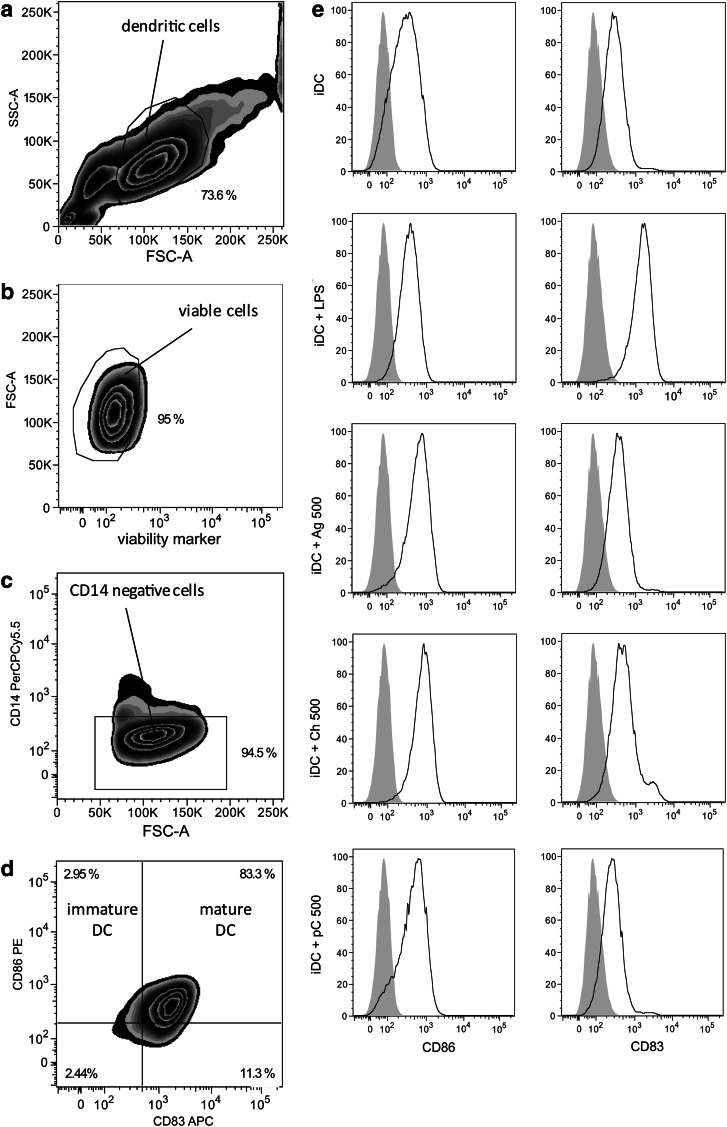
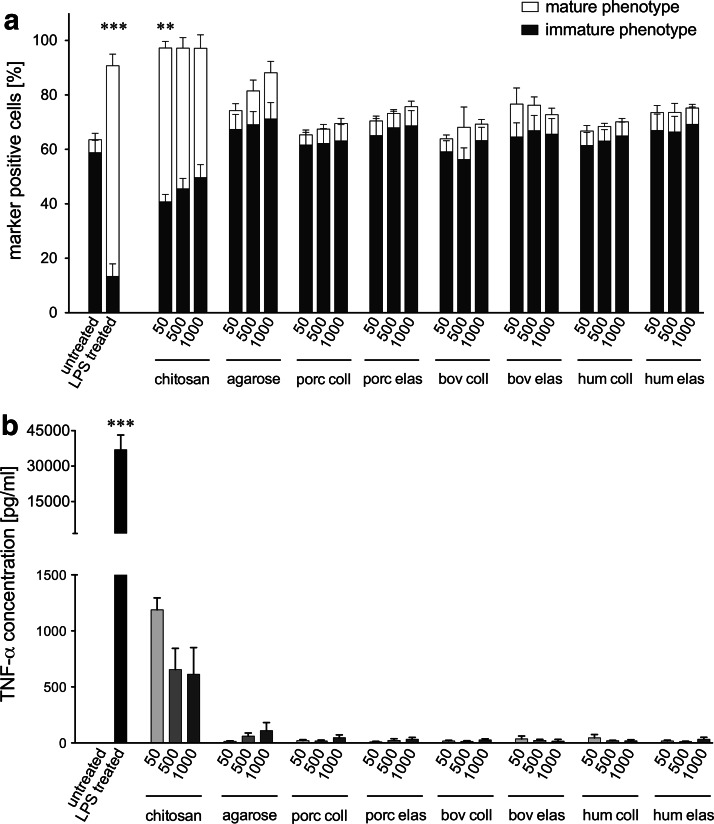
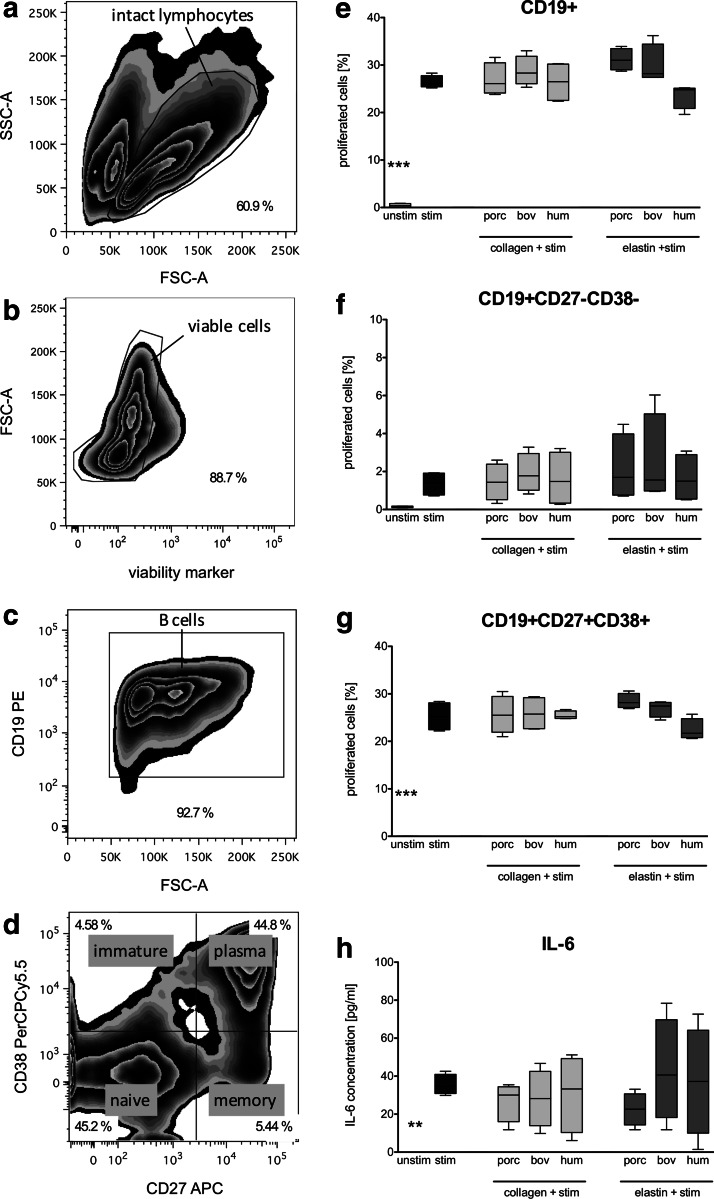
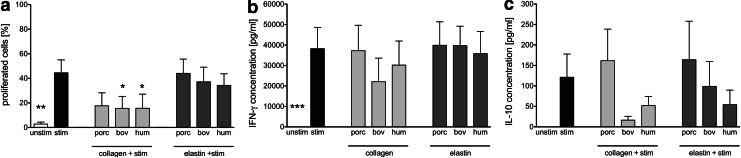
Novel tissue-engineering approaches for cardiovascular matrices based on xenogeneic extracellular matrix protein (ECMp) constituents require a detailed evaluation of their interaction with essential immune cell subsets playing a role in innate or adaptive immunity. Therefore, in this study, the effects of xenogeneic (porcine, bovine) collagen type I and elastin as the two main components of the heart valve ECM were analyzed in comparison to their human equivalents. First, their potential to induce maturation and cytokine secretion of human dendritic cells (DC) was tested by flow cytometry. Second, the influence on proliferation and cytokine release of purified human B and T cells was measured. We could demonstrate that xenogeneic collagen type I and elastin are not able to trigger the maturation of DC as verified by the lack of CD83 induction accompanied by a low tumor necrosis factor-α release. Moreover, both ECMp showed no effect on the proliferation and the interleukin-6 release of either unstimulated or prestimulated B cells. Additionally, anti-CD3-induced purified T cell proliferation and secretion of cytokines was not affected. All in vitro data verify the low immunogenicity of porcine and bovine collagen type I and elastin and favor their suitability for tissue-engineered scaffolds.
Figures
References
-
- Akhyari P. Minol P. Assmann A. Barth M. Kamiya H. Lichtenberg A. [Tissue engineering of heart valves] Chirurg. 2011;82:311. - PubMed
-
- Gilbert T.W. Sellaro T.L. Badylak S.F. Decellularization of tissues and organs. Biomaterials. 2006;27:3675. - PubMed
-
- Schoen F.J. Heart valve tissue engineering: quo vadis? Curr Opin Biotechnol. 2011;22:698. - PubMed
-
- Simon P. Kasimir M.T. Seebacher G. Weigel G. Ullrich R. Salzer-Muhar U. Rieder E. Wolner E. Early failure of the tissue engineered porcine heart valve SYNERGRAFT in pediatric patients. Eur J Cardiothorac Surg. 2003;23:1002. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
