

Reciprocal control of anaplerotic phosphoenolpyruvate carboxylase by in vivo monoubiquitination and phosphorylation in developing proteoid roots of phosphate-deficient harsh hakea
- PMID: 23407057
- PMCID: PMC3613444
- DOI: 10.1104/pp.112.213496
Reciprocal control of anaplerotic phosphoenolpyruvate carboxylase by in vivo monoubiquitination and phosphorylation in developing proteoid roots of phosphate-deficient harsh hakea
Abstract
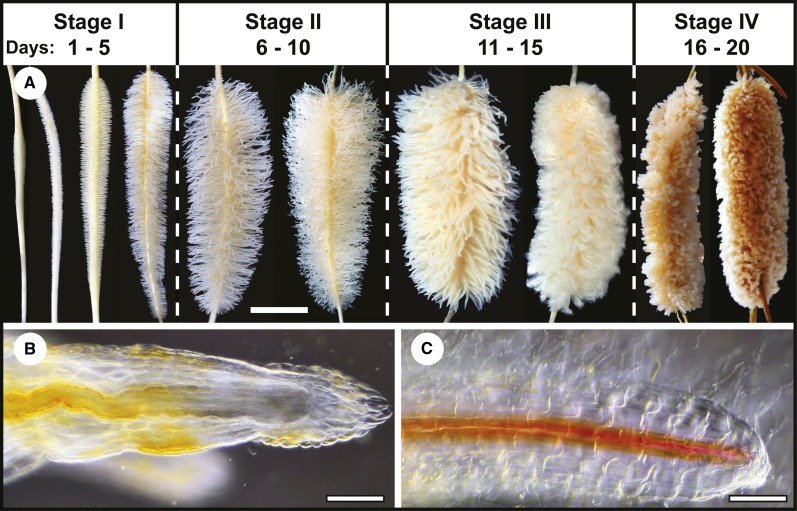
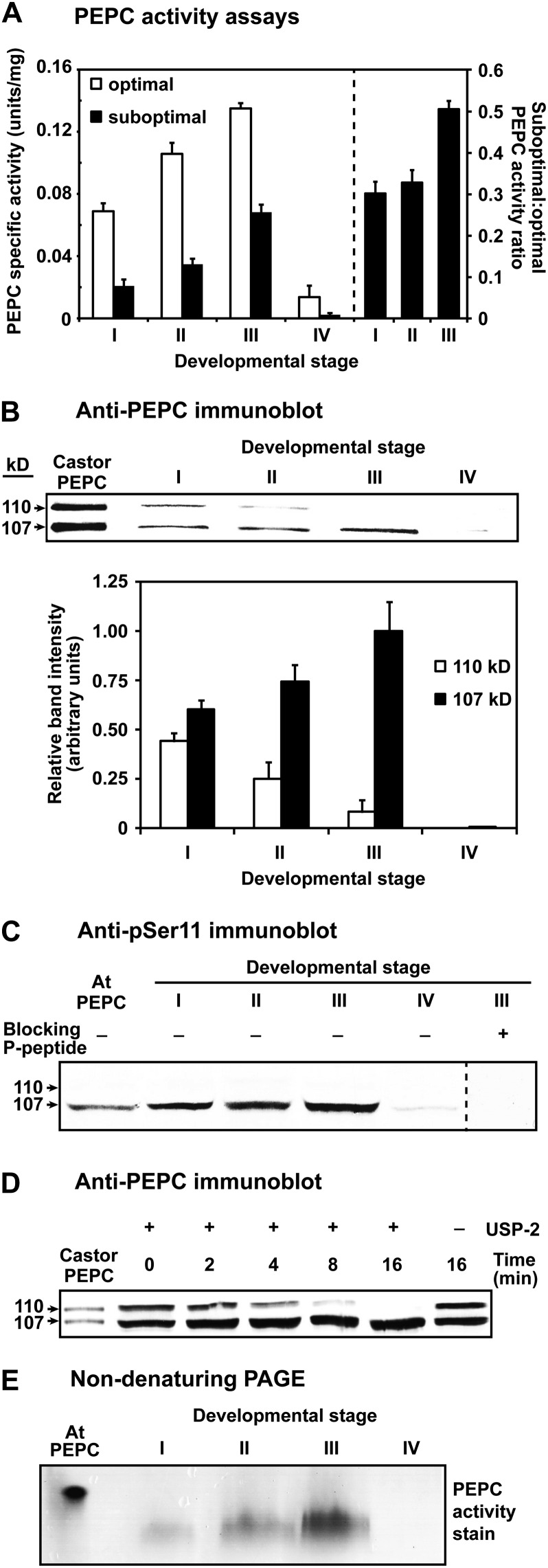
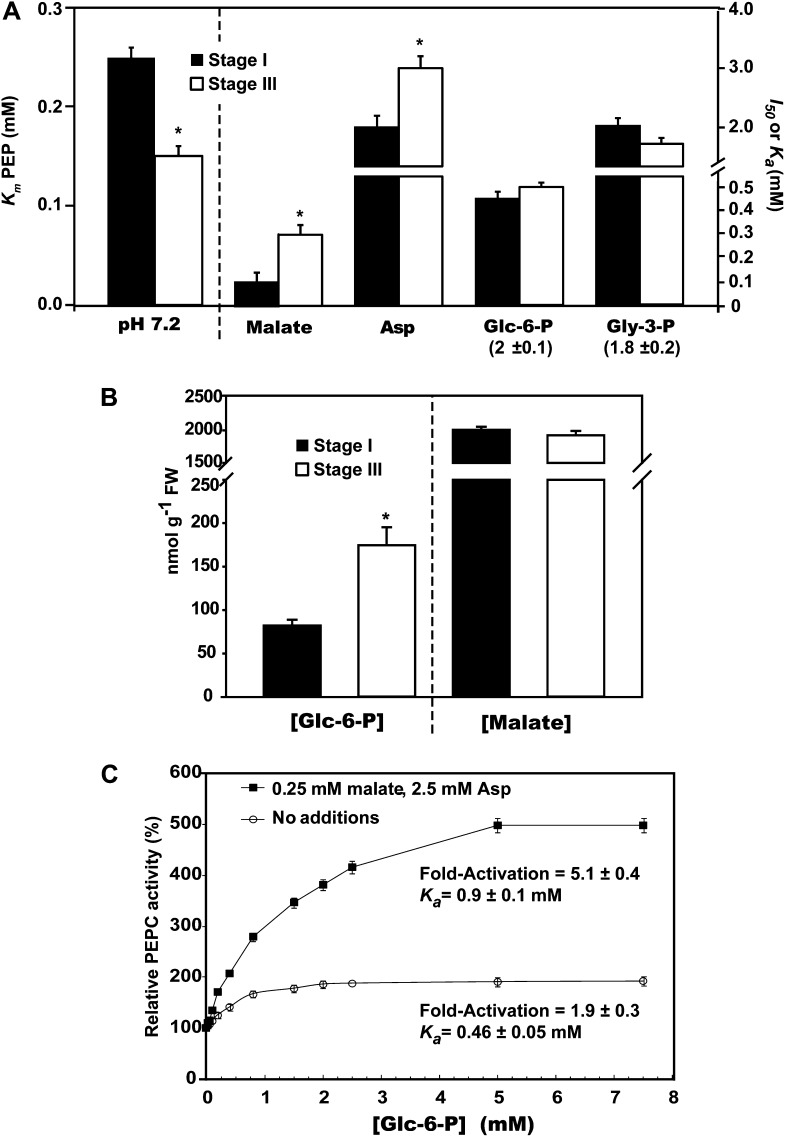
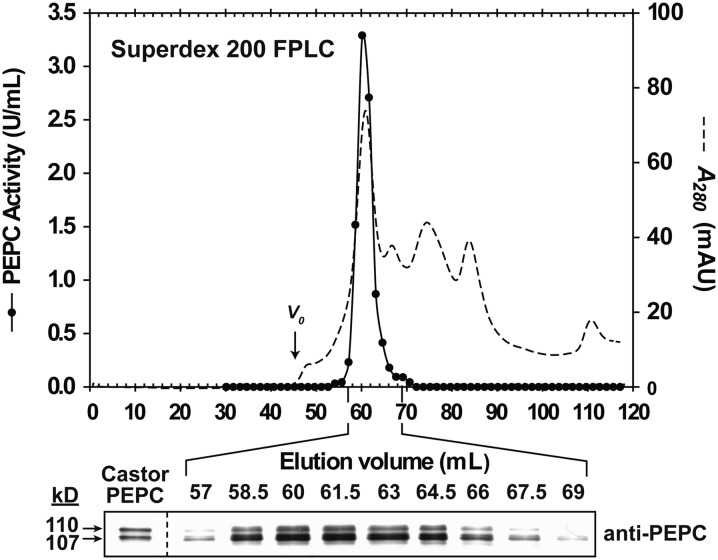
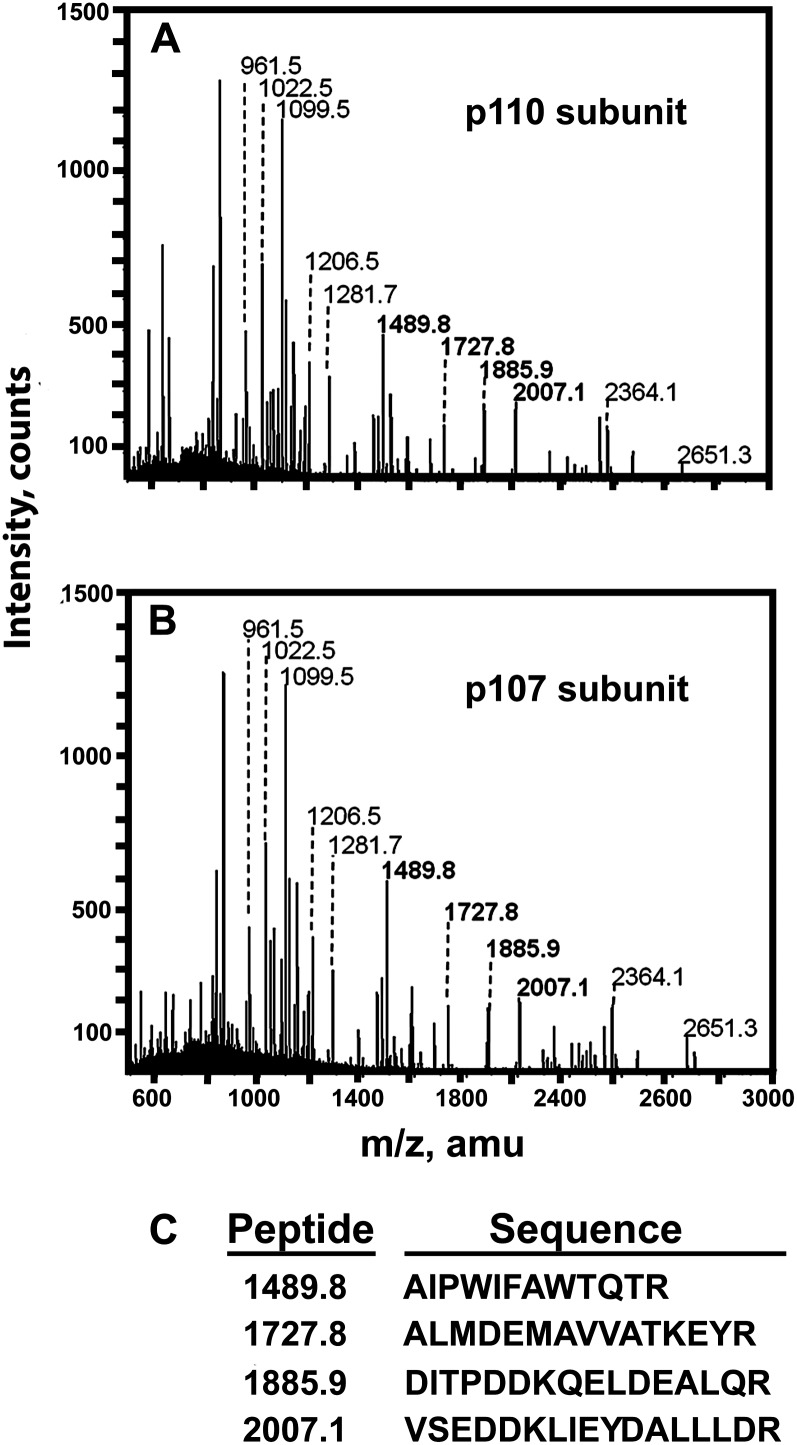
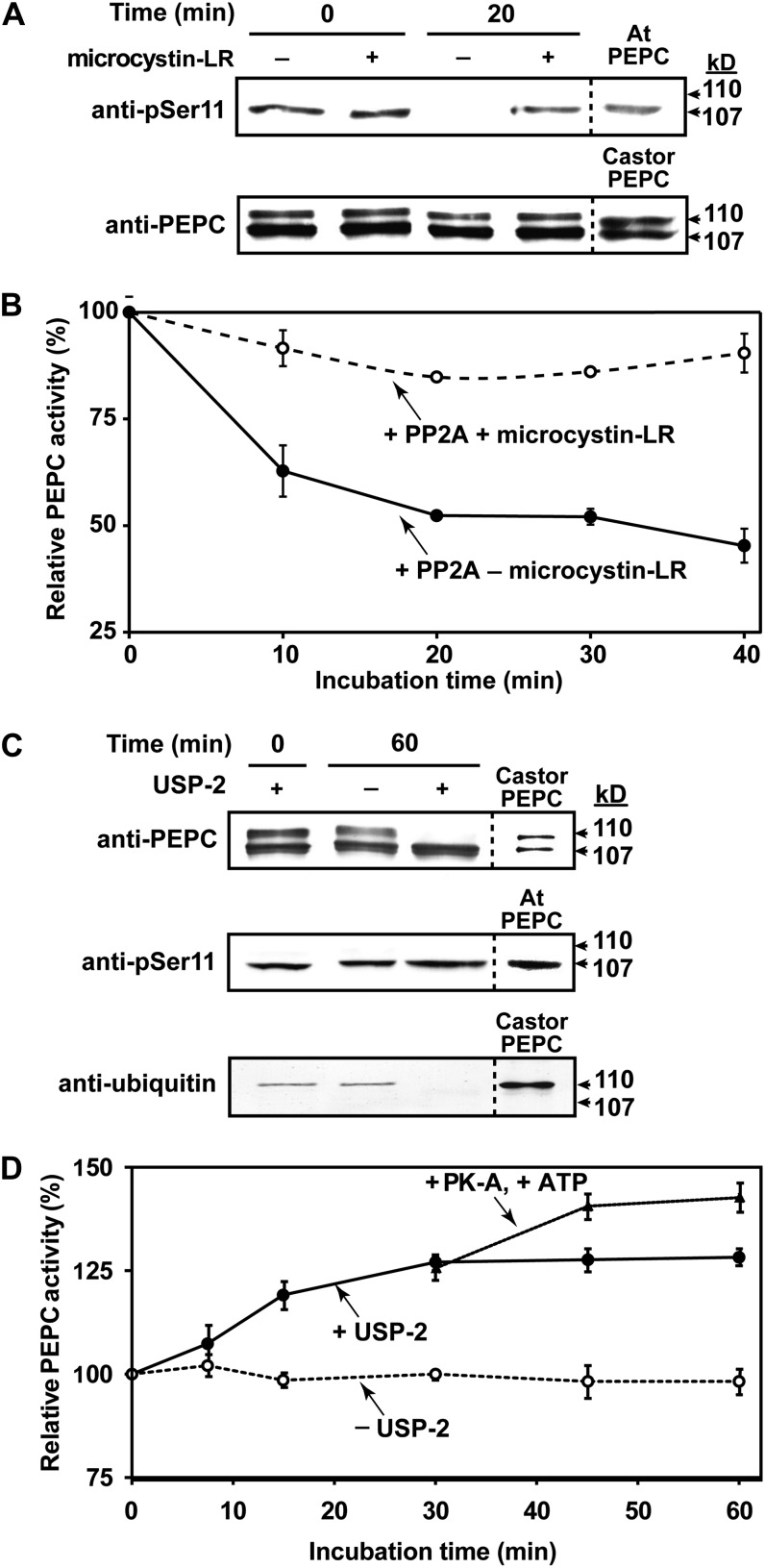
Accumulating evidence indicates important functions for phosphoenolpyruvate (PEP) carboxylase (PEPC) in inorganic phosphate (Pi)-starved plants. This includes controlling the production of organic acid anions (malate, citrate) that are excreted in copious amounts by proteoid roots of nonmycorrhizal species such as harsh hakea (Hakea prostrata). This, in turn, enhances the bioavailability of mineral-bound Pi by solubilizing Al(3+), Fe(3+), and Ca(2+) phosphates in the rhizosphere. Harsh hakea thrives in the nutrient-impoverished, ancient soils of southwestern Australia. Proteoid roots from Pi-starved harsh hakea were analyzed over 20 d of development to correlate changes in malate and citrate exudation with PEPC activity, posttranslational modifications (inhibitory monoubiquitination versus activatory phosphorylation), and kinetic/allosteric properties. Immature proteoid roots contained an equivalent ratio of monoubiquitinated 110-kD and phosphorylated 107-kD PEPC polypeptides (p110 and p107, respectively). PEPC purification, immunoblotting, and mass spectrometry indicated that p110 and p107 are subunits of a 430-kD heterotetramer and that they both originate from the same plant-type PEPC gene. Incubation with a deubiquitinating enzyme converted the p110:p107 PEPC heterotetramer of immature proteoid roots into a p107 homotetramer while significantly increasing the enzyme's activity under suboptimal but physiologically relevant assay conditions. Proteoid root maturation was paralleled by PEPC activation (e.g. reduced Km [PEP] coupled with elevated I50 [malate and Asp] values) via in vivo deubiquitination of p110 to p107, and subsequent phosphorylation of the deubiquitinated subunits. This novel mechanism of posttranslational control is hypothesized to contribute to the massive synthesis and excretion of organic acid anions that dominates the carbon metabolism of the mature proteoid roots.
Figures
References
-
- Brooks S. (1992) A simple computer program with statistical tests for the analysis of enzyme kinetics. Biotechniques 13: 906–911 - PubMed
-
- Gennidakis S, Rao S, Greenham K, Uhrig RG, O’Leary B, Snedden WA, Lu C, Plaxton WC. (2007) Bacterial- and plant-type phosphoenolpyruvate carboxylase polypeptides interact in the hetero-oligomeric Class-2 PEPC complex of developing castor oil seeds. Plant J 52: 839–849 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
