

Elasticity, structure, and relaxation of extended proteins under force
- PMID: 23407163
- PMCID: PMC3593838
- DOI: 10.1073/pnas.1300596110
Elasticity, structure, and relaxation of extended proteins under force
Abstract
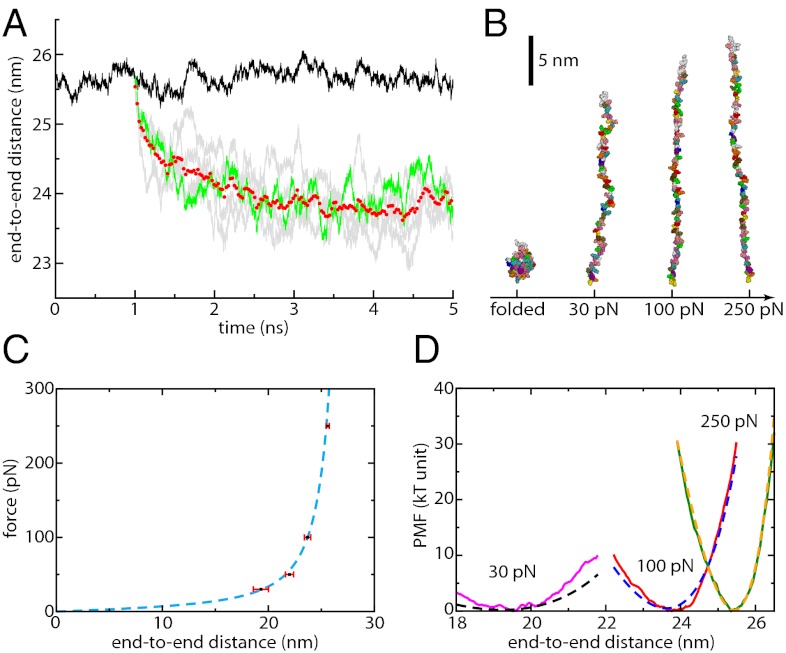
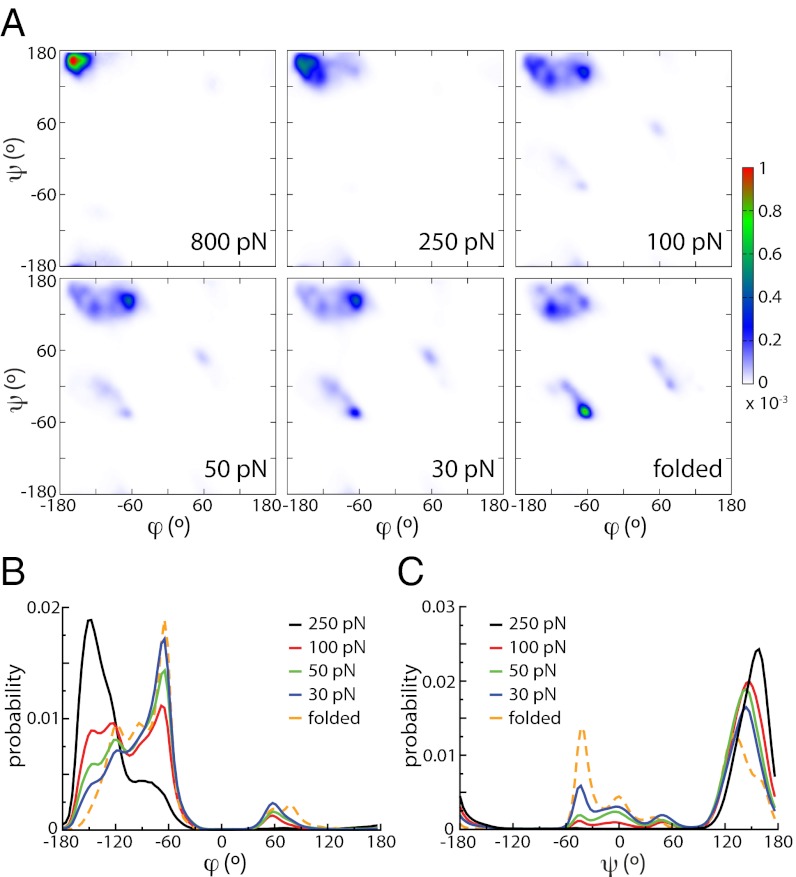
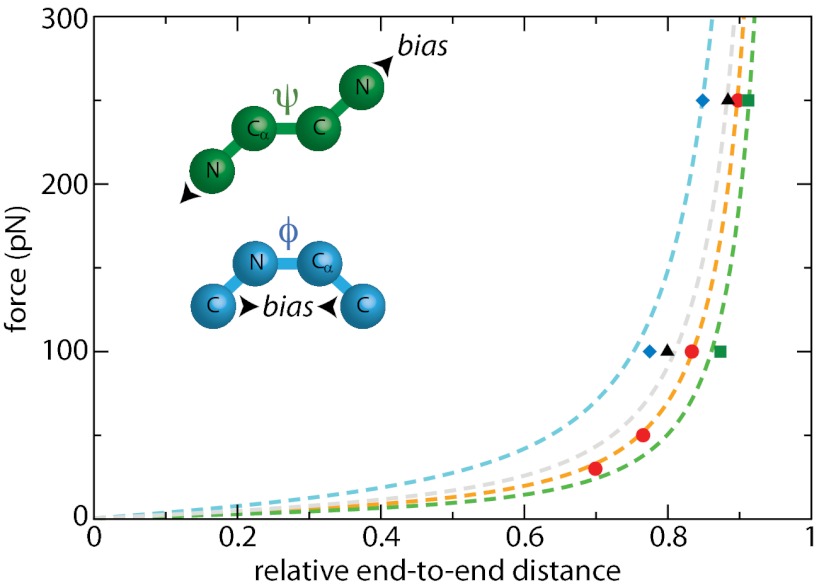
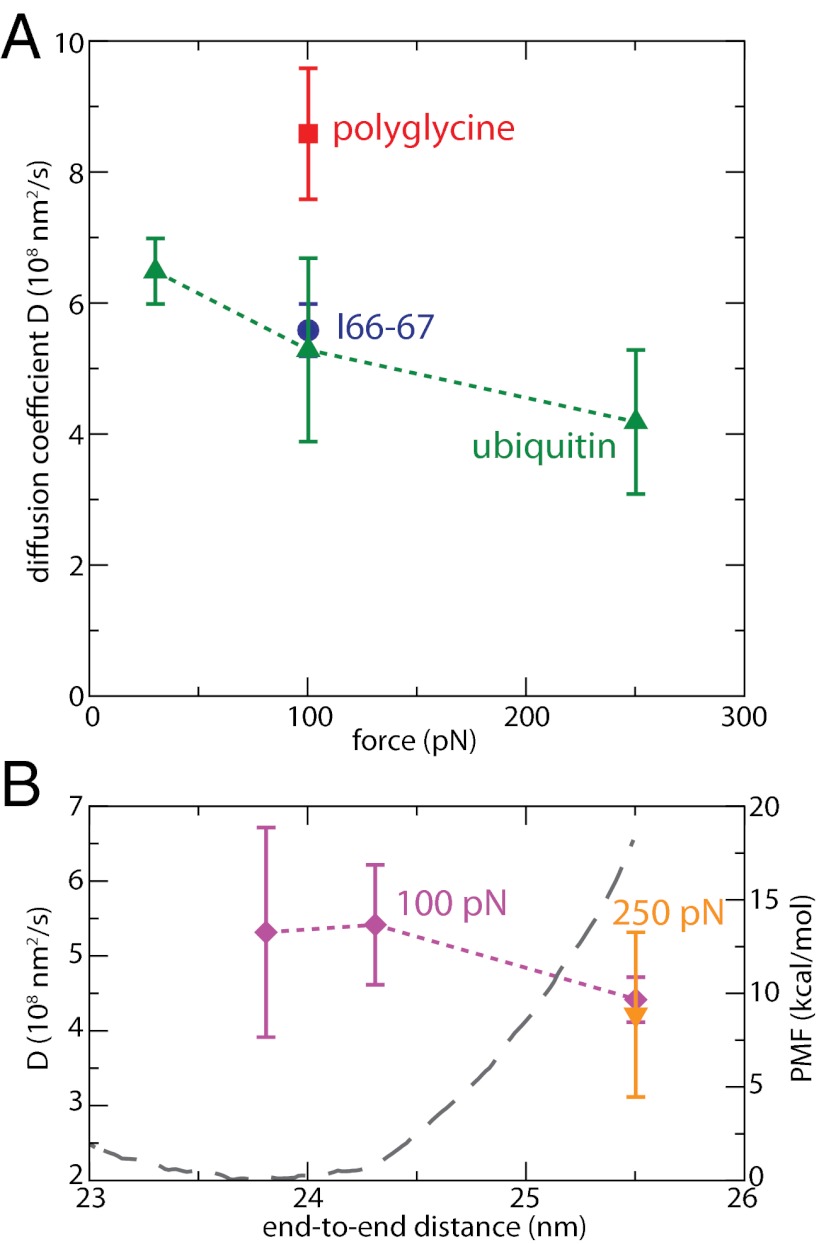
Force spectroscopies have emerged as a powerful and unprecedented tool to study and manipulate biomolecules directly at a molecular level. Usually, protein and DNA behavior under force is described within the framework of the worm-like chain (WLC) model for polymer elasticity. Although it has been surprisingly successful for the interpretation of experimental data, especially at high forces, the WLC model lacks structural and dynamical molecular details associated with protein relaxation under force that are key to the understanding of how force affects protein flexibility and reactivity. We use molecular dynamics simulations of ubiquitin to provide a deeper understanding of protein relaxation under force. We find that the WLC model successfully describes the simulations of ubiquitin, especially at higher forces, and we show how protein flexibility and persistence length, probed in the force regime of the experiments, are related to how specific classes of backbone dihedral angles respond to applied force. Although the WLC model is an average, backbone model, we show how the protein side chains affect the persistence length. Finally, we find that the diffusion coefficient of the protein's end-to-end distance is on the order of 10(8) nm(2)/s, is position and side-chain dependent, but is independent of the length and independent of the applied force, in contrast with other descriptions.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Bustamante C, Marko JF, Siggia ED, Smith S. Entropic elasticity of lambda-phage DNA. Science. 1994;265(5178):1599–1600. - PubMed
-
- Rief M, Gautel M, Oesterhelt F, Fernandez JM, Gaub HE. Reversible unfolding of individual titin immunoglobulin domains by AFM. Science. 1997;276(5315):1109–1112. - PubMed
-
- Merkel R, Nassoy P, Leung A, Ritchie K, Evans E. Energy landscapes of receptor-ligand bonds explored with dynamic force spectroscopy. Nature. 1999;397(6714):50–53. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
