

The structure of F₁-ATPase from Saccharomyces cerevisiae inhibited by its regulatory protein IF₁
- PMID: 23407639
- PMCID: PMC3603450
- DOI: 10.1098/rsob.120164
The structure of F₁-ATPase from Saccharomyces cerevisiae inhibited by its regulatory protein IF₁
Abstract
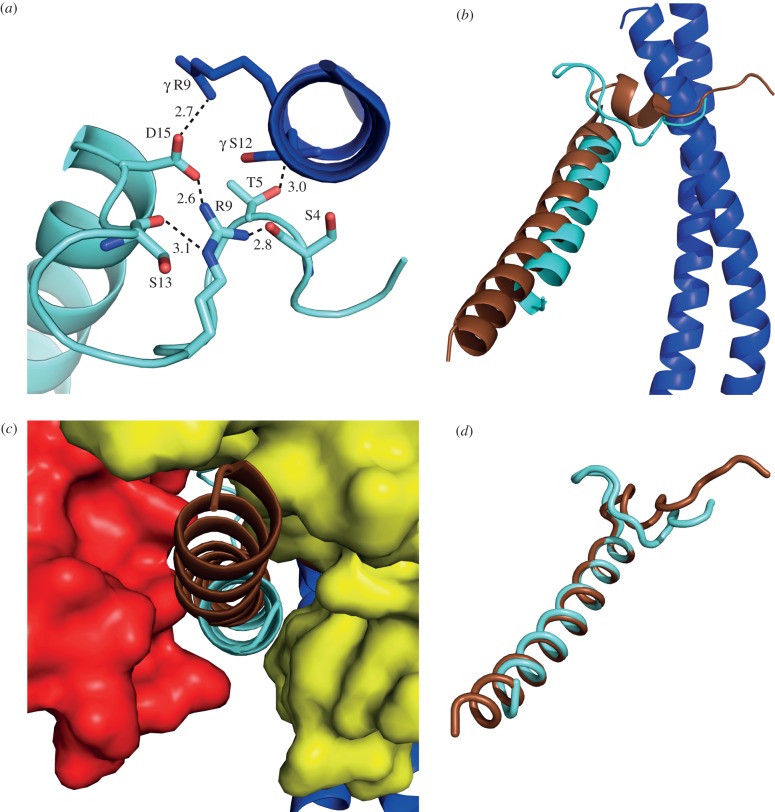
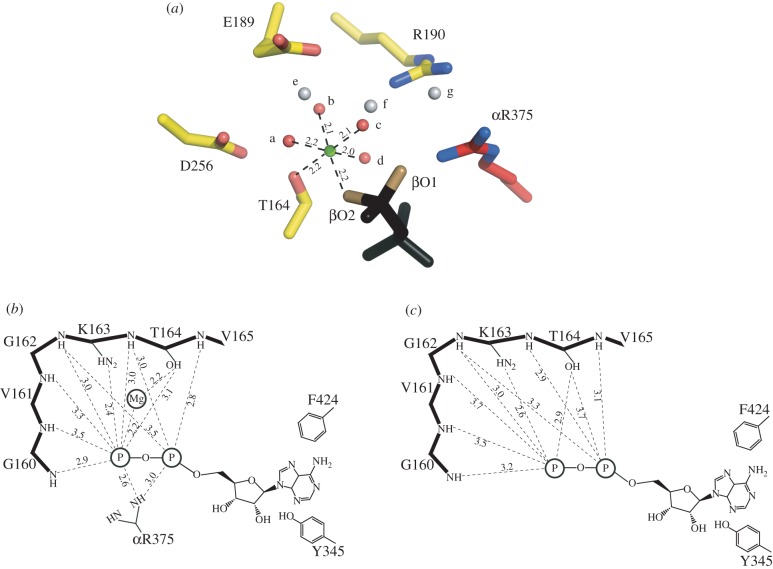
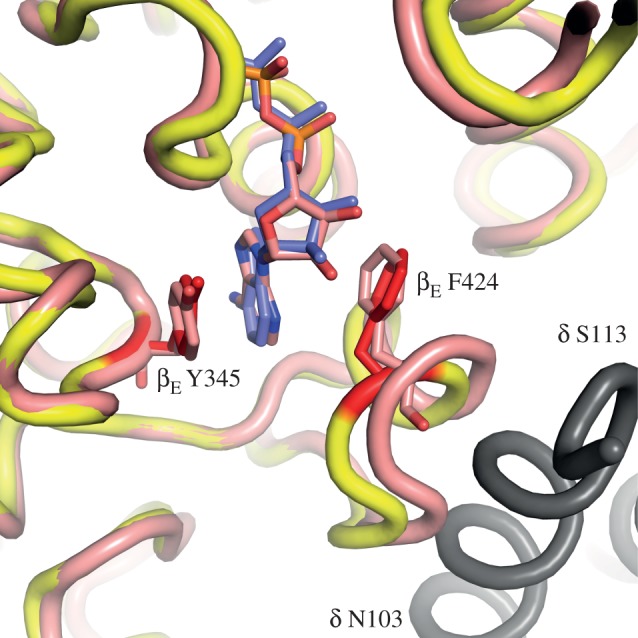
The structure of F₁-ATPase from Saccharomyces cerevisiae inhibited by the yeast IF₁ has been determined at 2.5 Å resolution. The inhibitory region of IF₁ from residues 1 to 36 is entrapped between the C-terminal domains of the α(DP)- and β(DP)-subunits in one of the three catalytic interfaces of the enzyme. Although the structure of the inhibited complex is similar to that of the bovine-inhibited complex, there are significant differences between the structures of the inhibitors and their detailed interactions with F₁-ATPase. However, the most significant difference is in the nucleotide occupancy of the catalytic β(E)-subunits. The nucleotide binding site in β(E)-subunit in the yeast complex contains an ADP molecule without an accompanying magnesium ion, whereas it is unoccupied in the bovine complex. Thus, the structure provides further evidence of sequential product release, with the phosphate and the magnesium ion released before the ADP molecule.
Figures




References
-
- Pullman ME, Penefsky H, Datta A, Racker E. 1960. Partial resolution of the enzymes catalysing oxidative phosphorylation. Purification and properties of soluble, dinitrophenol-stimulated adenosine triphosphatase. J. Biol. Chem. 235, 3322–3329 - PubMed
-
- Frangione B, Rosenwasser E, Penefsky HS, Pullman ME. 1981. Amino acid sequence of the protein inhibitor of mitochondrial adenosine triphosphatase. Proc. Natl Acad. Sci. USA 78, 7403–7407 10.1073/pnas.78.12.7403 (doi:10.1073/pnas.78.12.7403) - DOI - PMC - PubMed
-
- Walker JE, Gay NJ, Powell SJ, Kostina M, Dyer MR. 1987. ATP synthase from bovine mitochondria: sequences of imported precursors of oligomycin sensitivity conferral protein, factor 6, and adenosinetriphosphatase inhibitor protein. Biochemistry 26, 8613–8619 10.1021/bi00400a018 (doi:10.1021/bi00400a018) - DOI - PubMed
-
- Cabezon E, Butler PJ, Runswick MJ, Walker JE. 2000. Modulation of the oligomerization state of the bovine F1-ATPase inhibitor protein, IF1, by pH. J. Biol. Chem. 275, 25 460–25 464 10.1074/jbc.M003859200 (doi:10.1074/jbc.M003859200) - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases