

Processing of visual signals related to self-motion in the cerebellum of pigeons
- PMID: 23408161
- PMCID: PMC3569843
- DOI: 10.3389/fnbeh.2013.00004
Processing of visual signals related to self-motion in the cerebellum of pigeons
Abstract
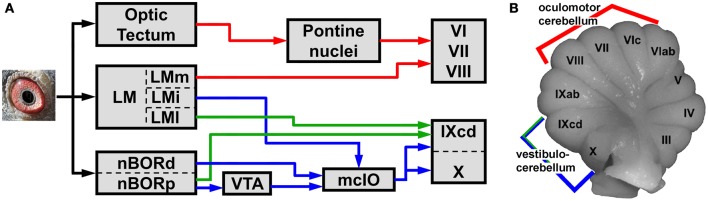
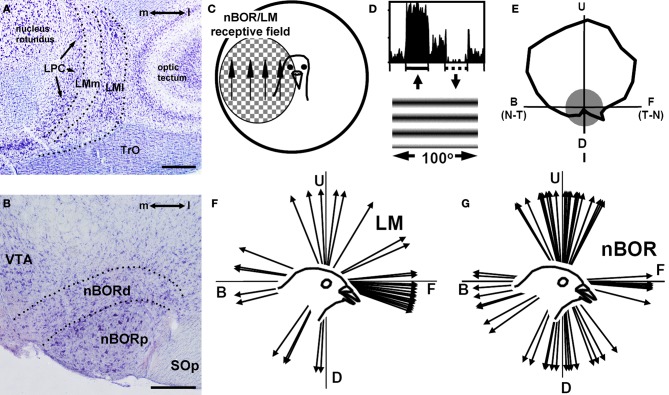
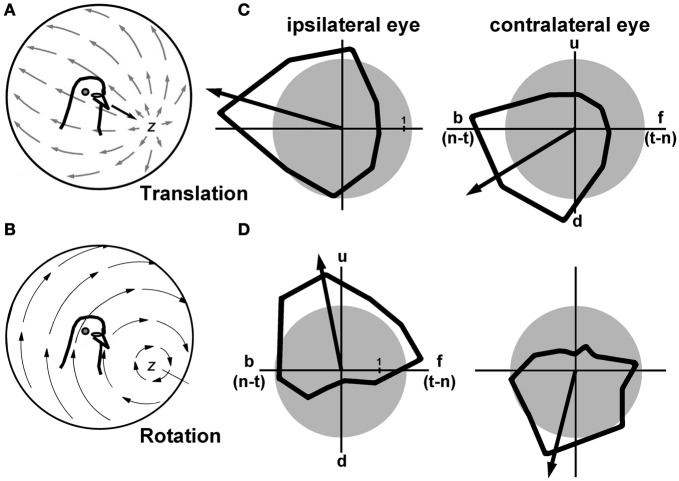
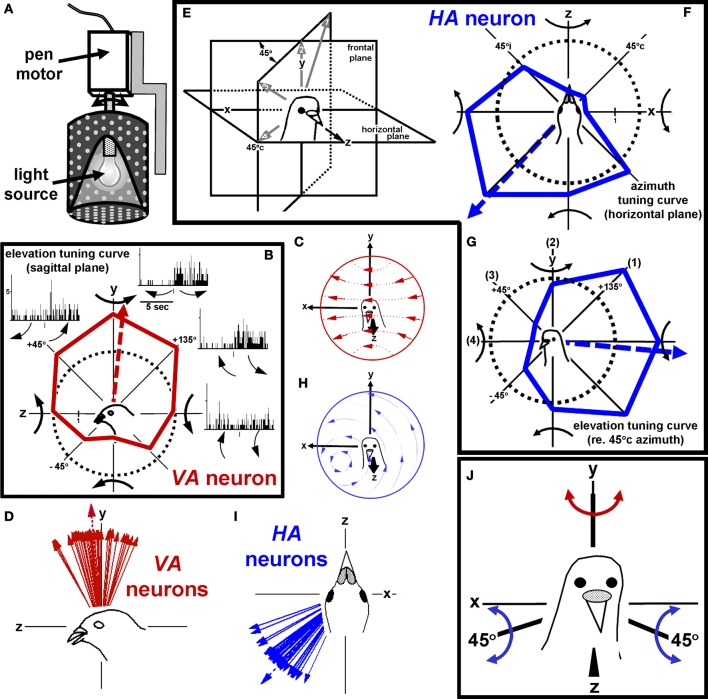
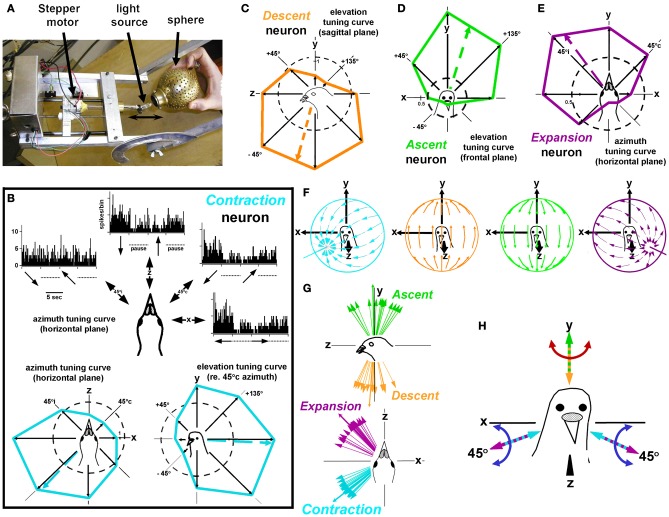
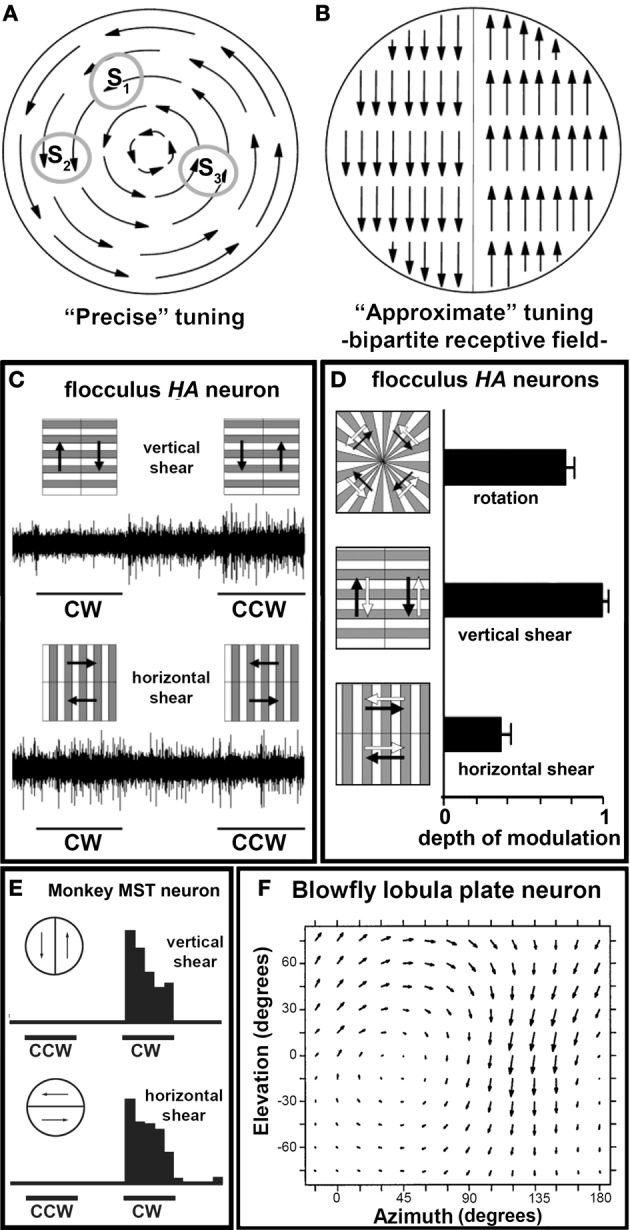
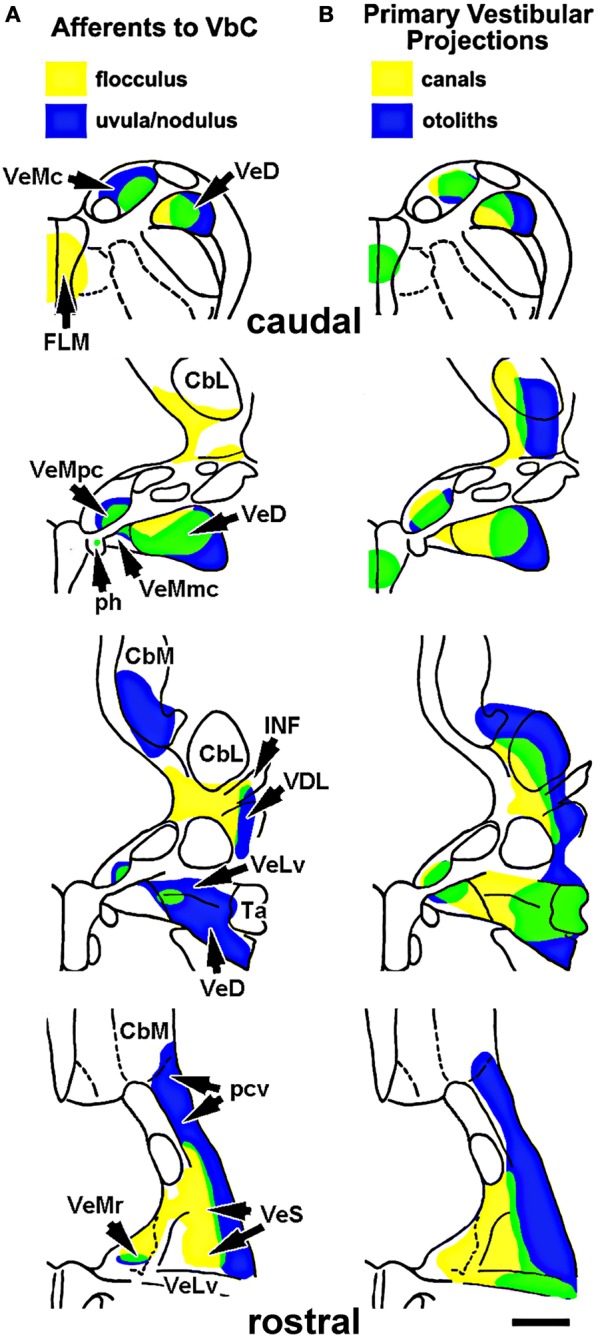
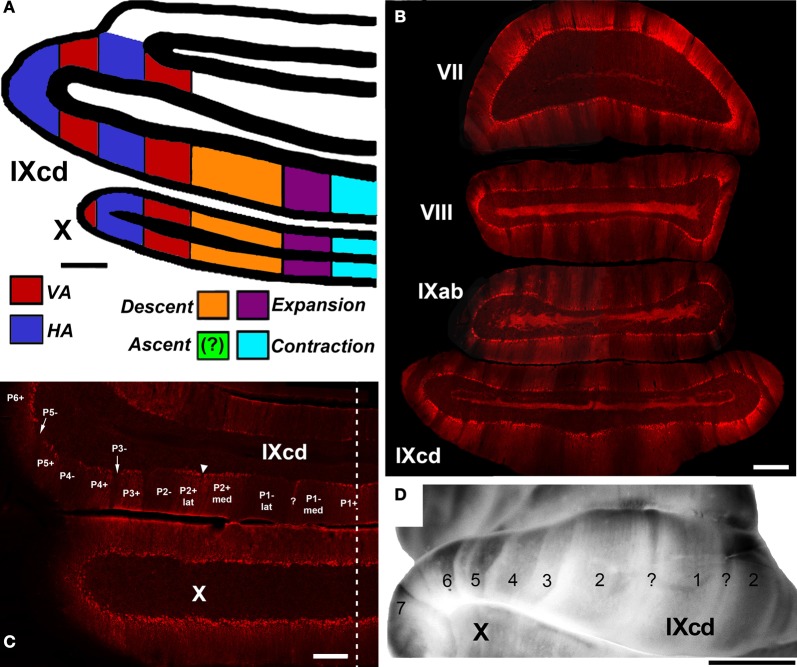
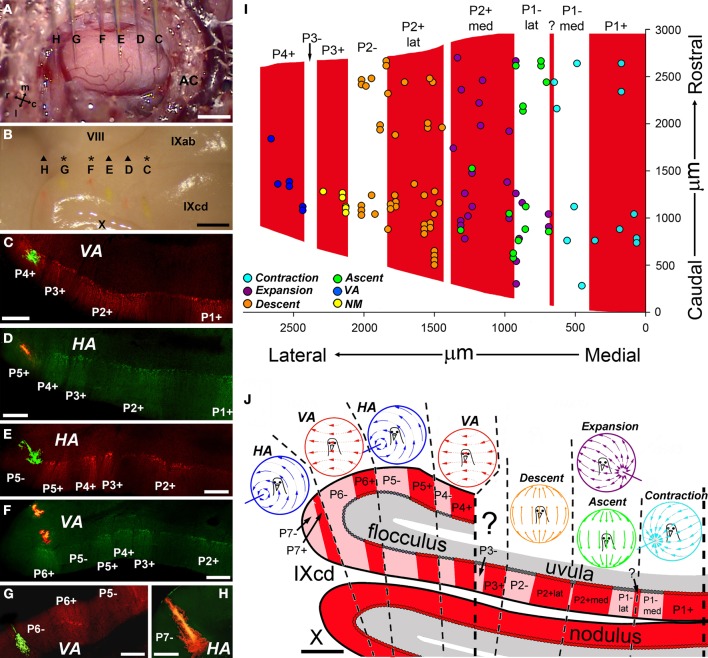
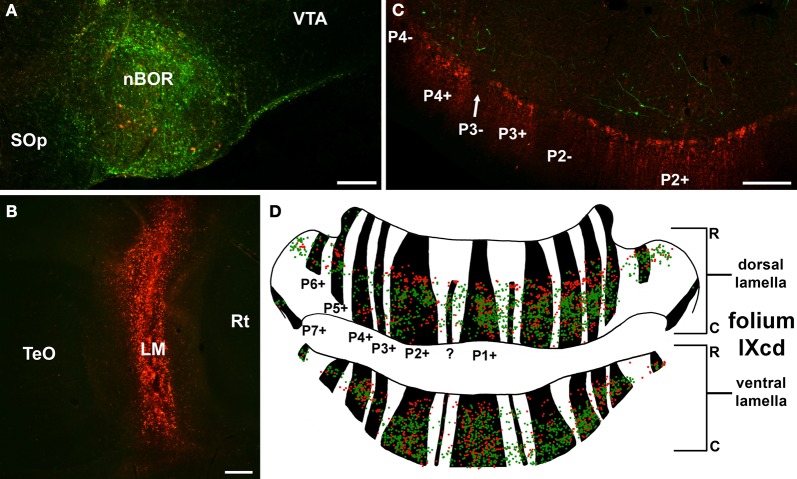
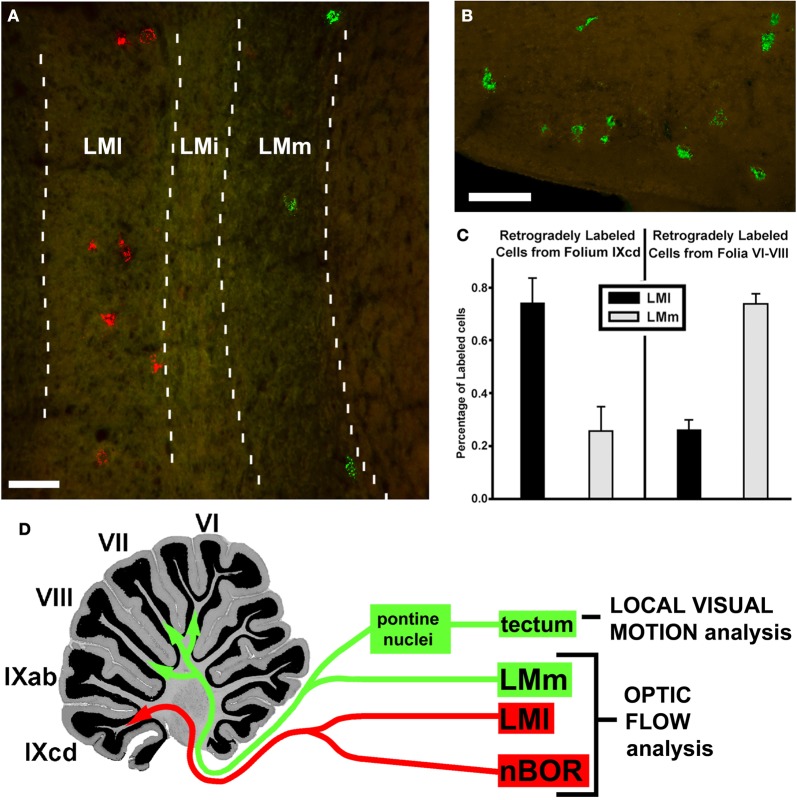
In this paper I describe the key features of optic flow processing in pigeons. Optic flow is the visual motion that occurs across the entire retina as a result of self-motion and is processed by subcortical visual pathways that project to the cerebellum. These pathways originate in two retinal-recipient nuclei, the nucleus of the basal optic root (nBOR) and the nucleus lentiformis mesencephali, which project to the vestibulocerebellum (VbC) (folia IXcd and X), directly as mossy fibers, and indirectly as climbing fibers from the inferior olive. Optic flow information is integrated with vestibular input in the VbC. There is a clear separation of function in the VbC: Purkinje cells in the flocculus process optic flow resulting from self-rotation, whereas Purkinje cells in the uvula/nodulus process optic flow resulting from self-translation. Furthermore, Purkinje cells with particular optic flow preferences are organized topographically into parasagittal "zones." These zones are correlated with expression of the isoenzyme aldolase C, also known as zebrin II (ZII). ZII expression is heterogeneous such that there are parasagittal stripes of Purkinje cells that have high expression (ZII+) alternating with stripes of Purkinje cells with low expression (ZII-). A functional zone spans a ZII± stripe pair. That is, each zone that contains Purkinje cells responsive to a particular pattern of optic flow is subdivided into a strip containing ZII+ Purkinje cells and a strip containing ZII- Purkinje cells. Additionally, there is optic flow input to folia VI-VIII of the cerebellum from lentiformis mesencephali. These folia also receive visual input from the tectofugal system via pontine nuclei. As the tectofugal system is involved in the analysis of local motion, there is integration of optic flow and local motion information in VI-VIII. This part of the cerebellum may be important for moving through a cluttered environment.
Keywords: accessory optic system; cerebellum; oculomotor cerebellum; optic flow; pretectum; vestibulocerebellum; zebrin.
Figures
References
-
- Ahn A. H., Dziennis S., Hawkes R., Herrup K. (1994). The cloning of zebrin II reveals its identity with aldolase C. Development 120, 2081–2090 - PubMed
-
- Andersson G., Oscarsson O. (1978). Climbing fiber microzones in cerebellar vermis and their projection to different groups of cells in the lateral vestibular nucleus. Exp. Brain Res. 32, 565–579 - PubMed
-
- Arends J. J. A., Voogd J. (1989). Topographic aspects of the olivocerebellar system in the pigeon. Exp. Brain Res. 17(Suppl.), 52–57
-
- Barmack N. H., Shojaku H. (1992). Representation of a postural coordinate system in the nodulus of the rabbit cerebellum by vestibular climbing fiber signals, in Vestibular Control of Eye, Head and Body Movements, eds Shimazu H., Shinoda Y. (Tokyo; Japan Scientific Societies Press; Basel: Karger; ), 331–338
-
- Brauth S. E., Karten H. J. (1977). Direct accessory optic projections to vestibulo-cerebellum– possible channel for oculomotor control-systems. Exp. Brain Res. 28, 73–84 - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources