

Membrane topology and function of dengue virus NS2A protein
- PMID: 23408612
- PMCID: PMC3624351
- DOI: 10.1128/JVI.02424-12
Membrane topology and function of dengue virus NS2A protein
Abstract
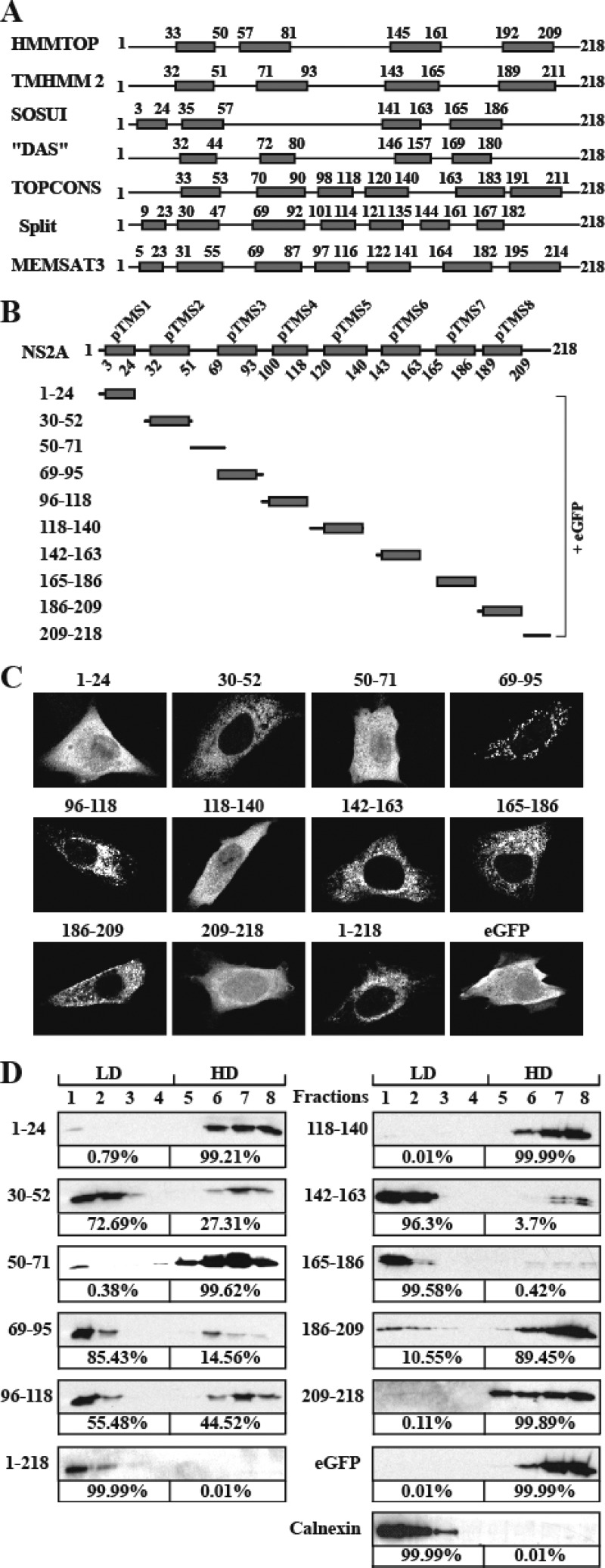
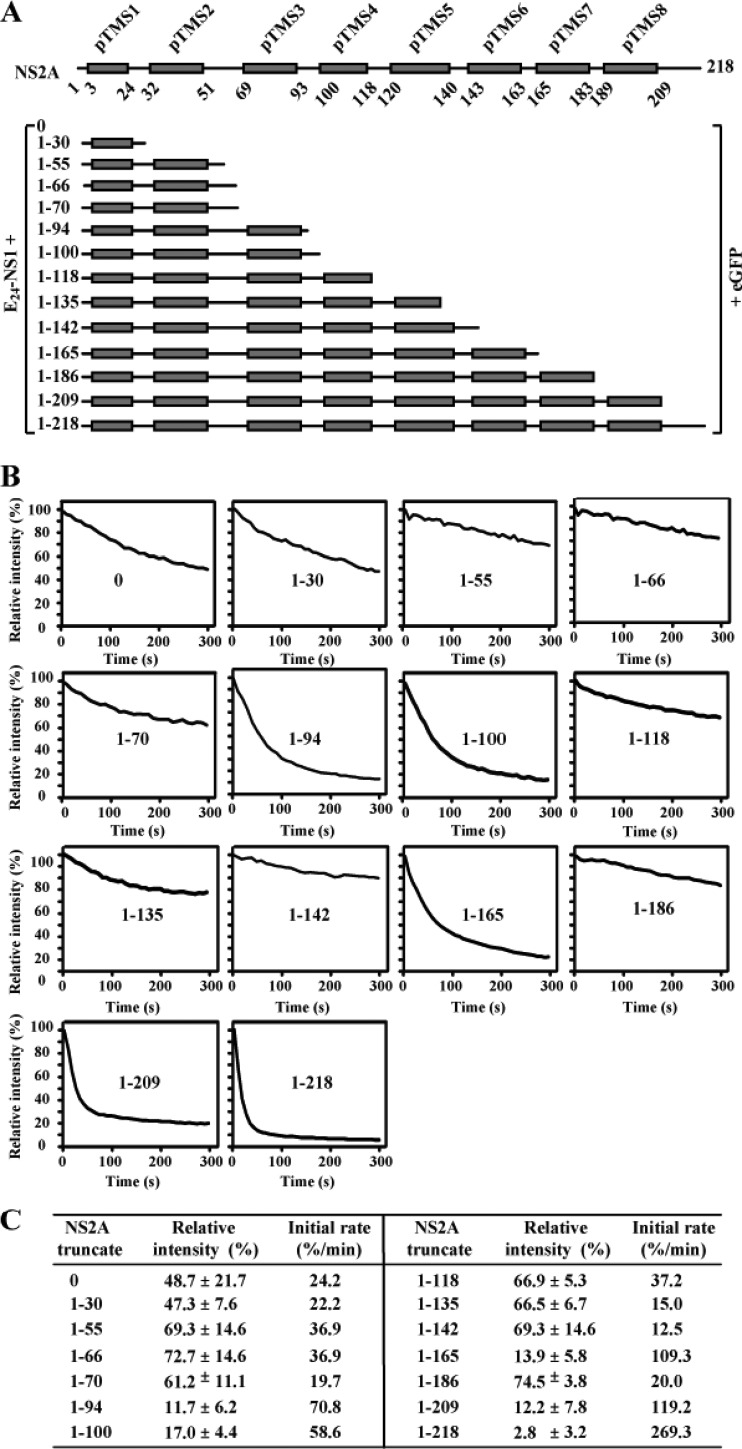
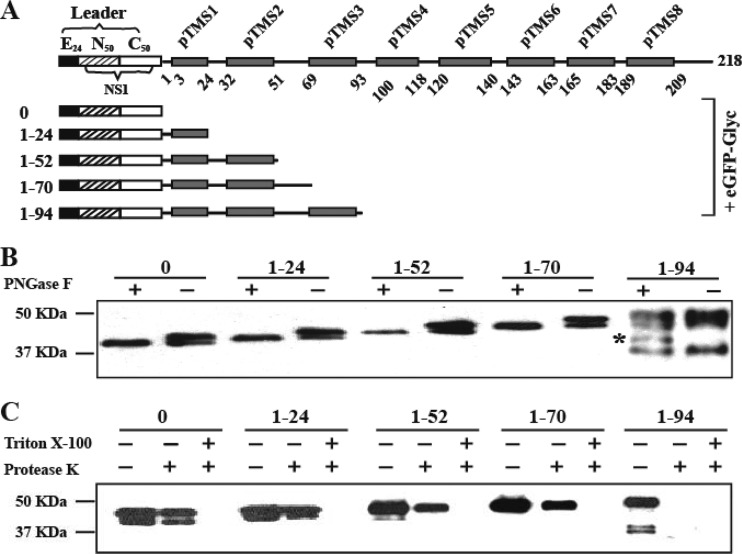
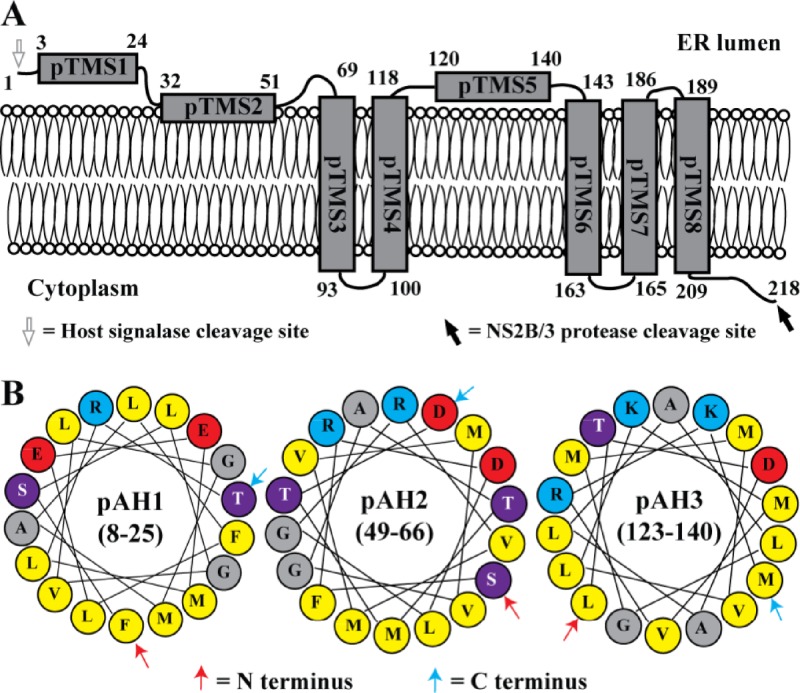
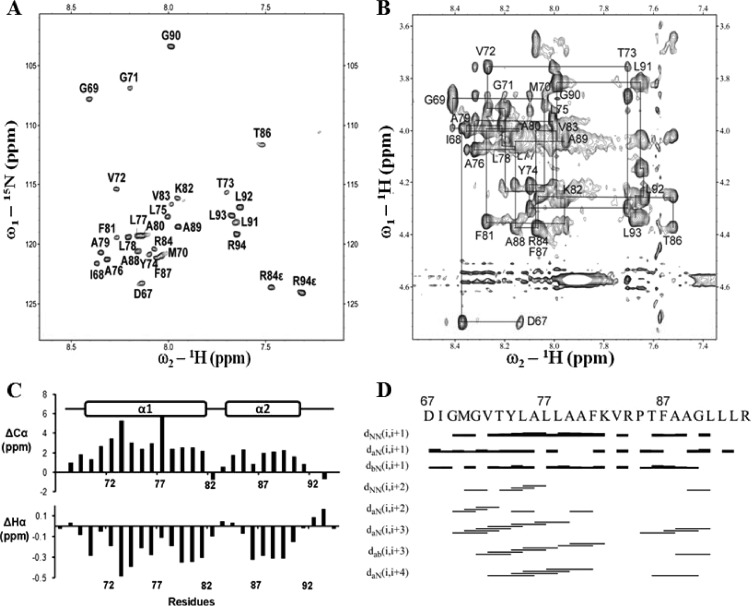
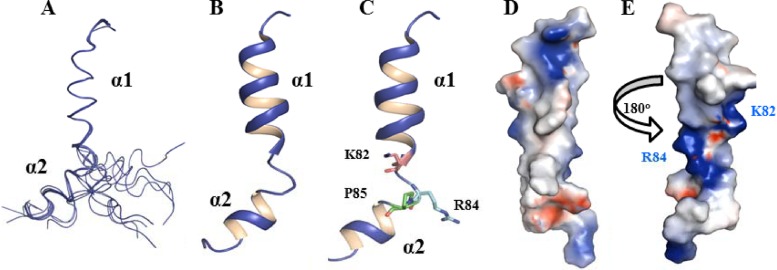
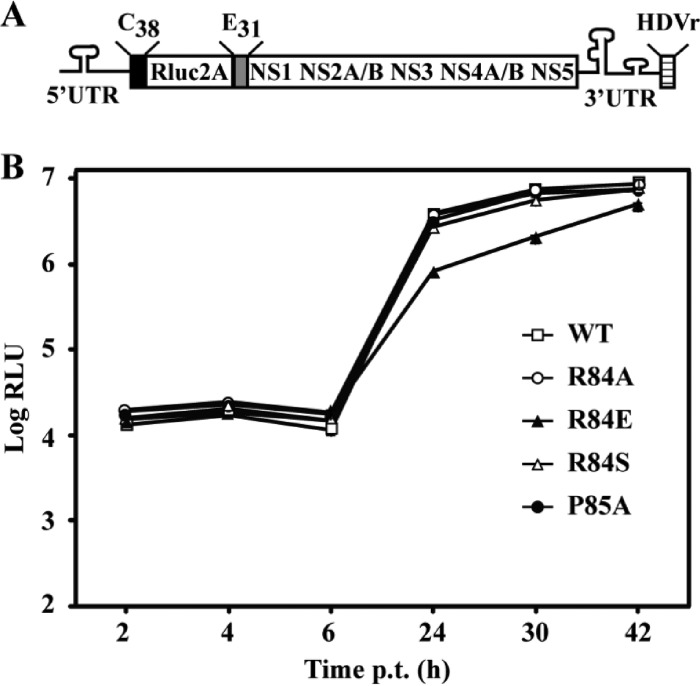
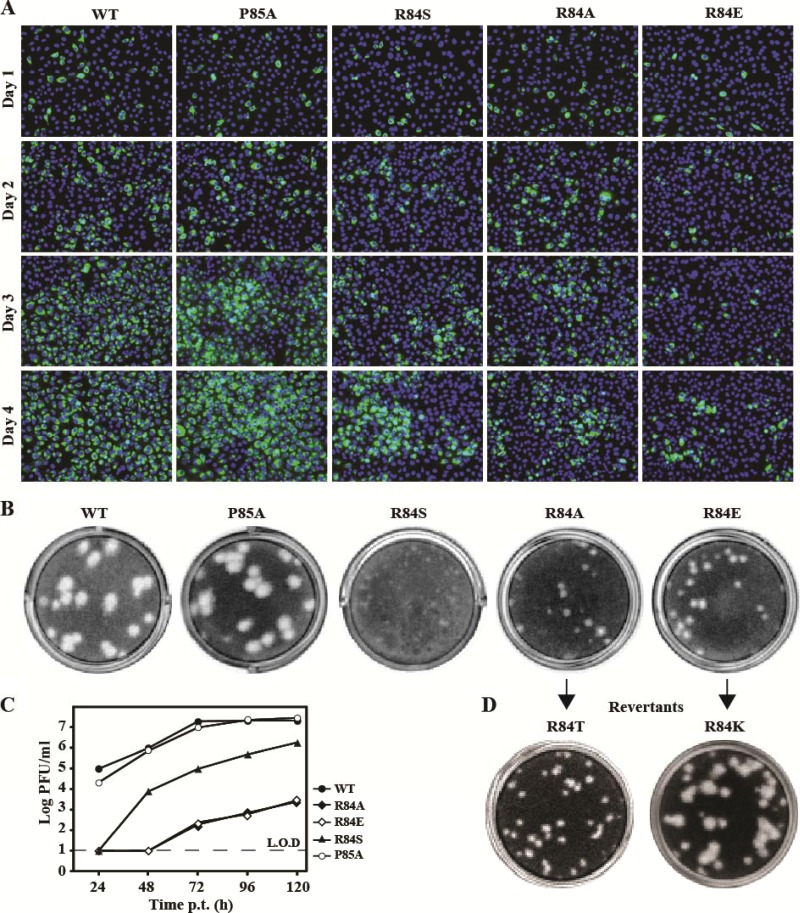
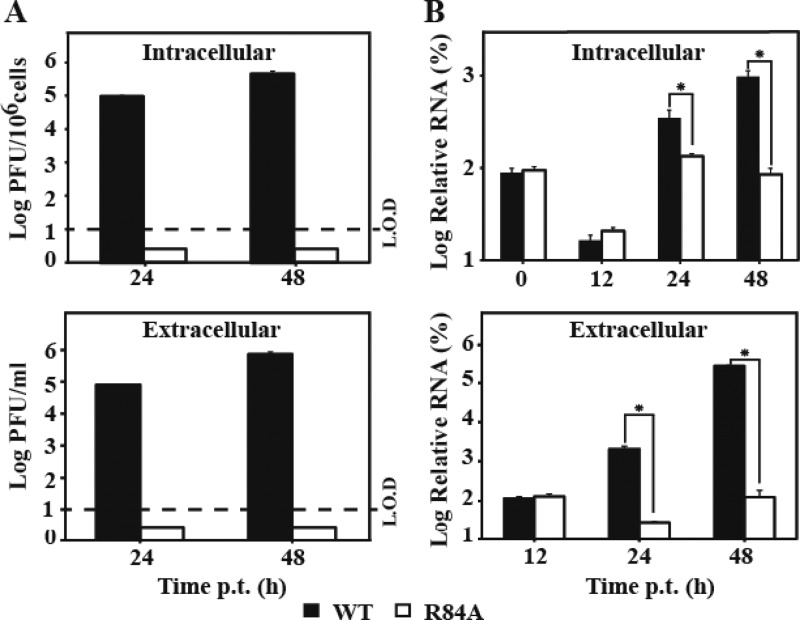
Flavivirus nonstructural protein 2A (NS2A) is a component of the viral replication complex that functions in virion assembly and antagonizes the host immune response. Although flavivirus NS2A is known to associate with the endoplasmic reticulum (ER) membrane, the detailed topology of this protein has not been determined. Here we report the first topology model of flavivirus NS2A on the ER membrane. Using dengue virus (DENV) NS2A as a model, we show that (i) the N-terminal 68 amino acids are located in the ER lumen, with one segment (amino acids 30 to 52) that interacts with ER membrane without traversing the lipid bilayer; (ii) amino acids 69 to 209 form five transmembrane segments, each of which integrally spans the ER membrane; and (iii) the C-terminal tail (amino acids 210 to 218) is located in the cytosol. Nuclear magnetic resonance (NMR) structural analysis showed that the first membrane-spanning segment (amino acids 69 to 93) consists of two helices separated by a "helix breaker." The helix breaker is formed by amino acid P85 and one positively charged residue, R84. Functional analysis using replicon and genome-length RNAs of DENV-2 indicates that P85 is not important for viral replication. However, when R84 was replaced with E, the mutation attenuated both viral RNA synthesis and virus production. Remarkably, an R84A mutation did not affect viral RNA synthesis but blocked intracellular formation of infectious virions. Collectively, the mutagenesis results demonstrate that NS2A functions in both DENV RNA synthesis and virion assembly/maturation. The topology model of DENV NS2A provides a good starting point for studying how flavivirus NS2A modulates viral replication and evasion of host immune response.
Figures
References
-
- Lindenbach BD, Thiel H-J, Rice CM. 2007. Flaviviridae: the viruses and their replication, p 1101–1152 In Knipe DM, Howley PM. (ed), Fields virology, 5th ed, vol 1 Lippincott William & Wilkins, Philadelphia, PA
-
- Cleaves GR, Dubin DT. 1979. Methylation status of intracellular dengue type 2 40 S RNA. Virology 96:159–165 - PubMed
-
- Mackenzie JM, Khromykh AA, Jones MK, Westaway EG. 1998. Subcellular localization and some biochemical properties of the flavivirus Kunjin nonstructural proteins NS2A and NS4A. Virology 245:203–215 - PubMed
-
- Avirutnan P, Punyadee N, Noisakran S, Komoltri C, Thiemmeca S, et al. 2006. Vascular leakage in severe dengue virus infections: a potential role for the nonstructural viral protein NS1 and complement. J. Infect. Dis. 193:1078–1088 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
