

Nucleozin targets cytoplasmic trafficking of viral ribonucleoprotein-Rab11 complexes in influenza A virus infection
- PMID: 23408618
- PMCID: PMC3624347
- DOI: 10.1128/JVI.03123-12
Nucleozin targets cytoplasmic trafficking of viral ribonucleoprotein-Rab11 complexes in influenza A virus infection
Abstract
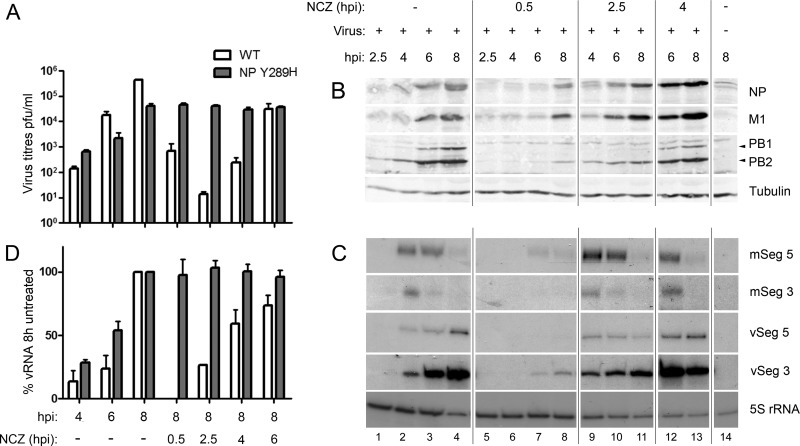
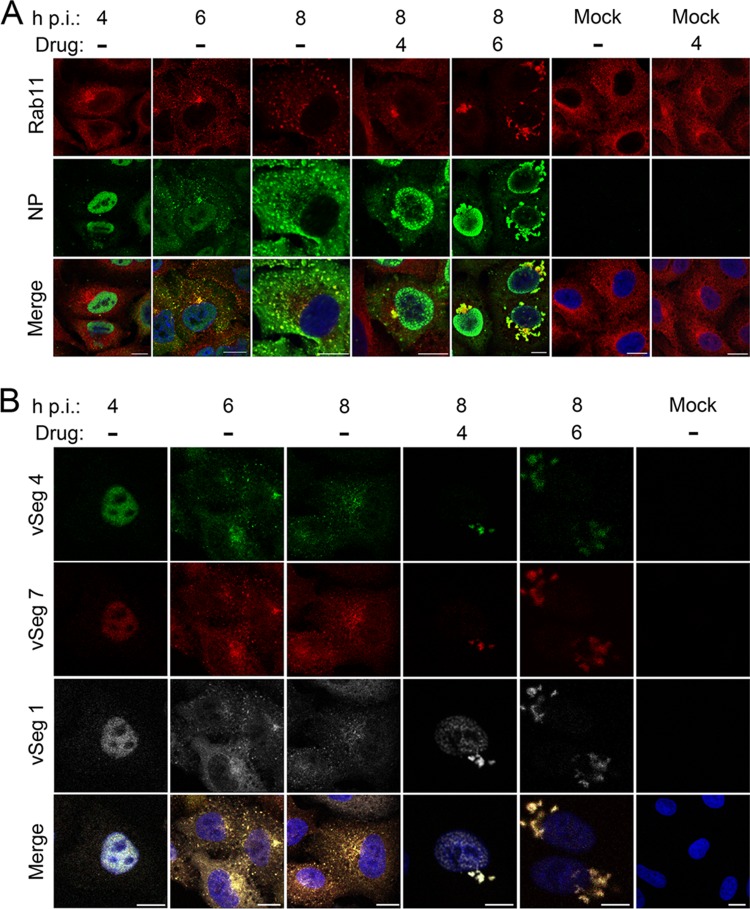
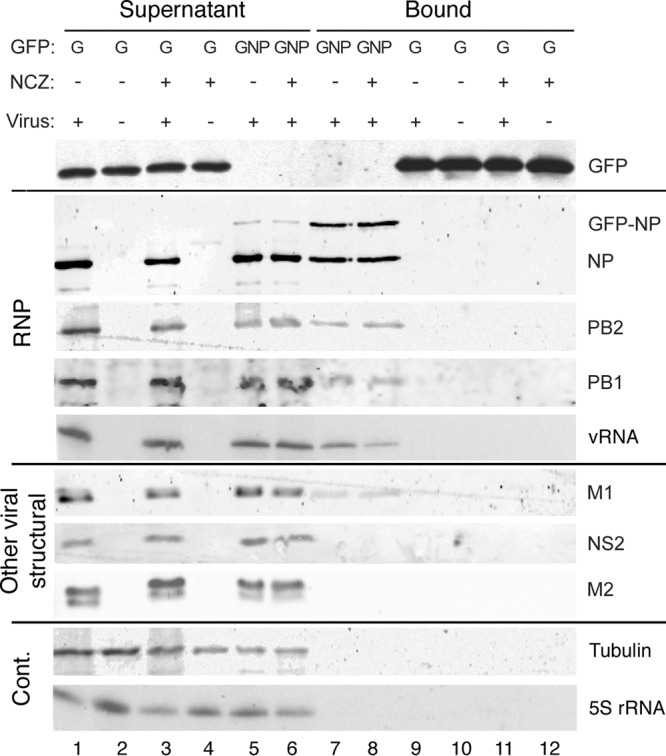
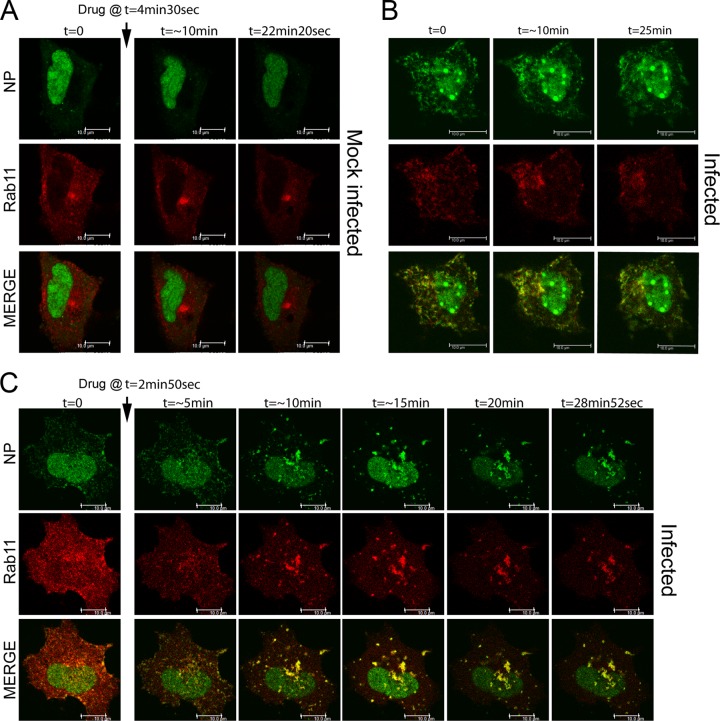
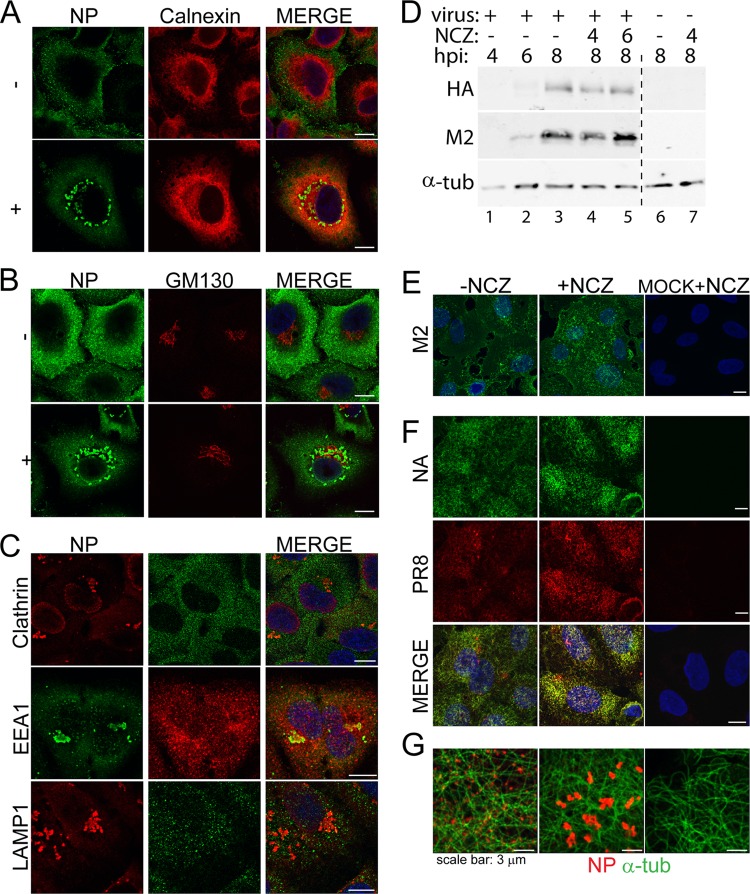
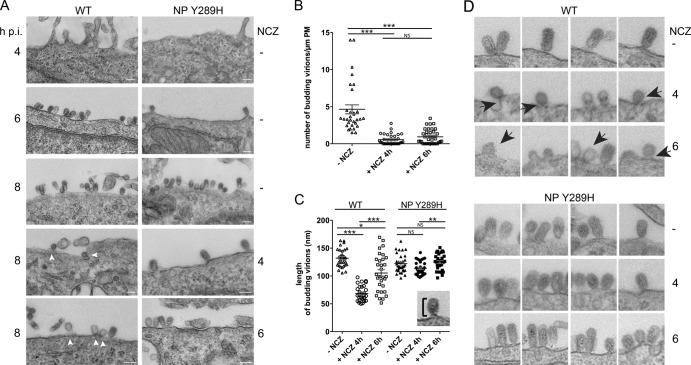
Novel antivirals are needed to supplement existing control strategies for influenza A virus (IAV). A promising new class of drug, exemplified by the compound nucleozin, has recently been identified that targets the viral nucleoprotein (NP). These inhibitors are thought to act as "molecular staples" that stabilize interactions between NP monomers, promoting the formation of nonfunctional aggregates. Here we detail the inhibitory mechanism of nucleozin, finding that the drug has both early- and late-acting effects on the IAV life cycle. When present at the start of infection, it inhibited viral RNA and protein synthesis. However, when added at later time points, it still potently blocked the production of infectious progeny but without affecting viral macromolecular synthesis. Instead, nucleozin blocked the cytoplasmic trafficking of ribonucleoproteins (RNPs) that had undergone nuclear export, promoting the formation of large perinuclear aggregates of RNPs along with cellular Rab11. This effect led to the production of much reduced amounts of often markedly smaller virus particles. We conclude that the primary target of nucleozin is the viral RNP, not NP, and this work also provides proof of the principle that IAV replication can be effectively inhibited by blocking cytoplasmic trafficking of the viral genome.
Figures
References
-
- Jagger BW, Wise HM, Kash JC, Walters KA, Wills NM, Xiao YL, Dunfee RL, Schwartzman LM, Ozinsky A, Bell GL, Dalton RM, Lo A, Efstathiou S, Atkins JF, Firth AE, Taubenberger JK, Digard P. 2012. An overlapping protein-coding region in influenza A virus segment 3 modulates the host response. Science 337:199–204 - PMC - PubMed
-
- Wise HM, Hutchinson EC, Jagger BW, Stuart AD, Kang ZH, Robb N, Schwartzman LM, Kash JC, Fodor E, Firth AE, Gog JR, Taubenberger JK, Digard P. 2012. Identification of a novel splice variant form of the influenza A virus M2 ion channel with an antigenically distinct ectodomain. PLoS Pathog. 8:e1002998 doi:10.1371/journal.ppat.1002998 - DOI - PMC - PubMed
-
- Portela A, Digard P. 2002. The influenza virus nucleoprotein: a multifunctional RNA-binding protein pivotal to virus replication. J. Gen. Virol. 83:723–734 - PubMed
-
- Boulo S, Akarsu H, Ruigrok RW, Baudin F. 2007. Nuclear traffic of influenza virus proteins and ribonucleoprotein complexes. Virus Res. 124:12–21 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
