

Viral capsid proteins are segregated in structural fold space
- PMID: 23408879
- PMCID: PMC3567143
- DOI: 10.1371/journal.pcbi.1002905
Viral capsid proteins are segregated in structural fold space
Abstract
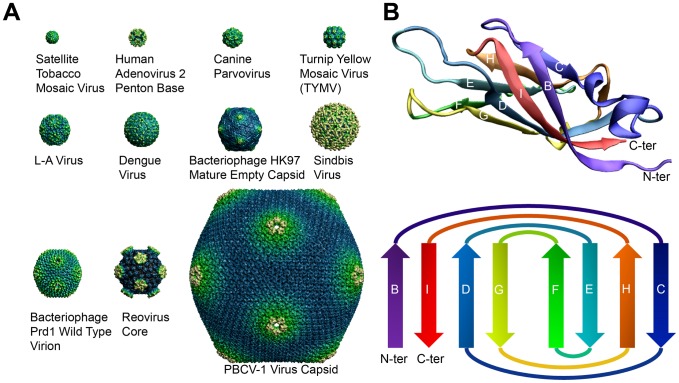
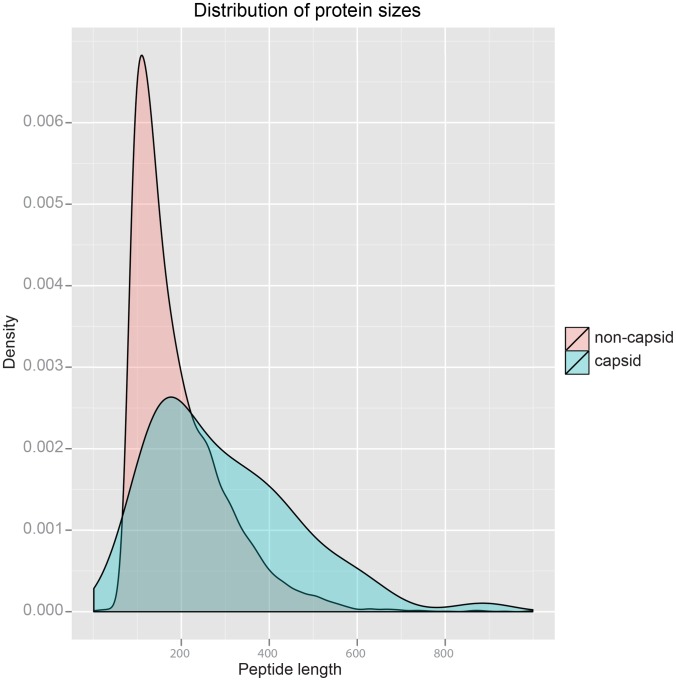
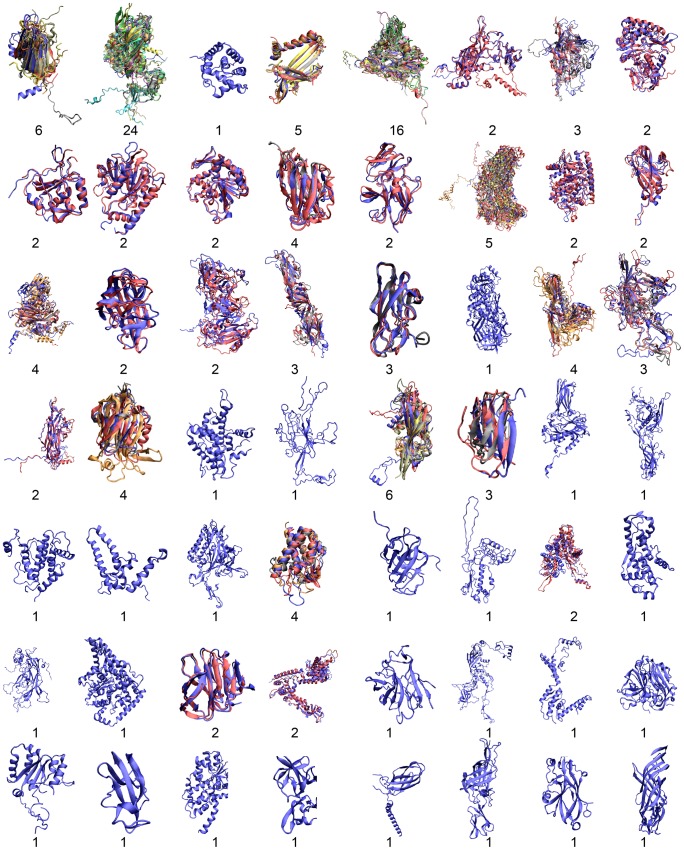
Viral capsid proteins assemble into large, symmetrical architectures that are not found in complexes formed by their cellular counterparts. Given the prevalence of the signature jelly-roll topology in viral capsid proteins, we are interested in whether these functionally unique capsid proteins are also structurally unique in terms of folds. To explore this question, we applied a structure-alignment based clustering of all protein chains in VIPERdb filtered at 40% sequence identity to identify distinct capsid folds, and compared the cluster medoids with a non-redundant subset of protein domains in the SCOP database, not including the viral capsid entries. This comparison, using Template Modeling (TM)-score, identified 2078 structural "relatives" of capsid proteins from the non-capsid set, covering altogether 210 folds following the definition in SCOP. The statistical significance of the 210 folds shared by two sets of the same sizes, estimated from 10,000 permutation tests, is less than 0.0001, which is an upper bound on the p-value. We thus conclude that viral capsid proteins are segregated in structural fold space. Our result provides novel insight on how structural folds of capsid proteins, as opposed to their surface chemistry, might be constrained during evolution by requirement of the assembled cage-like architecture. Also importantly, our work highlights a guiding principle for virus-based nanoplatform design in a wide range of biomedical applications and materials science.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures




Similar articles
-
Protein-Protein Interfaces in Viral Capsids Are Structurally Unique.J Mol Biol. 2015 Nov 6;427(22):3613-3624. doi: 10.1016/j.jmb.2015.09.008. Epub 2015 Sep 12. J Mol Biol. 2015. PMID: 26375252 Free PMC article.
-
The structure of the bacteriophage PRD1 spike sheds light on the evolution of viral capsid architecture.Mol Cell. 2005 Apr 15;18(2):161-70. doi: 10.1016/j.molcel.2005.03.019. Mol Cell. 2005. PMID: 15837420
-
Nucleic and Amino Acid Sequences Support Structure-Based Viral Classification.J Virol. 2017 Mar 29;91(8):e02275-16. doi: 10.1128/JVI.02275-16. Print 2017 Apr 15. J Virol. 2017. PMID: 28122979 Free PMC article.
-
Chrysovirus structure: repeated helical core as evidence of gene duplication.Adv Virus Res. 2013;86:87-108. doi: 10.1016/B978-0-12-394315-6.00004-0. Adv Virus Res. 2013. PMID: 23498904 Review.
-
IgStrand: A universal residue numbering scheme for the immunoglobulin-fold (Ig-fold) to study Ig-proteomes and Ig-interactomes.PLoS Comput Biol. 2025 Apr 14;21(4):e1012813. doi: 10.1371/journal.pcbi.1012813. eCollection 2025 Apr. PLoS Comput Biol. 2025. PMID: 40228037 Free PMC article. Review.
Cited by
-
Structural insights into conformational switching in latency-associated peptide between transforming growth factor β-1 bound and unbound states.IUCrJ. 2020 Feb 6;7(Pt 2):238-252. doi: 10.1107/S205225251901707X. eCollection 2020 Mar 1. IUCrJ. 2020. PMID: 32148852 Free PMC article.
-
Vaccine development and therapeutic design for 2019-nCoV/SARS-CoV-2: Challenges and chances.J Cell Physiol. 2020 Dec;235(12):9098-9109. doi: 10.1002/jcp.29771. Epub 2020 Jun 18. J Cell Physiol. 2020. PMID: 32557648 Free PMC article. Review.
-
Cyanophages as an important factor in the early evolution of oxygenic photosynthesis.Sci Rep. 2022 Nov 29;12(1):20581. doi: 10.1038/s41598-022-24795-1. Sci Rep. 2022. PMID: 36446879 Free PMC article.
-
Unmasking of the von Willebrand A-domain surface adhesin CglB at bacterial focal adhesions mediates myxobacterial gliding motility.Sci Adv. 2023 Feb 22;9(8):eabq0619. doi: 10.1126/sciadv.abq0619. Epub 2023 Feb 22. Sci Adv. 2023. PMID: 36812310 Free PMC article.
-
Host cell stress response as a predictor of COVID-19 infectivity and disease progression.Front Mol Biosci. 2022 Aug 11;9:938099. doi: 10.3389/fmolb.2022.938099. eCollection 2022. Front Mol Biosci. 2022. PMID: 36032680 Free PMC article. Review.
References
-
- Rux JJ, Burnett RM (1998) Spherical viruses. Current Opinion in Structural Biology 8: 142–149. - PubMed
-
- Harrison SC (2001) The familiar and the unexpected in structures of icosahedral viruses. Current Opinion in Structural Biology 11: 195–199. - PubMed
-
- Caspar DL, Klug A (1962) Physical principles in the construction of regular viruses. Cold Spring Harb Symp Quant Biol 27: 1–24. - PubMed
-
- Johnson JE, Chiu W (2000) Structures of virus and virus-like particles. Curr Opin Struct Biol 10: 229–235. - PubMed
-
- Filman DJ, Hogle JM (1991) Virology. Architecture with a difference. Nature 351: 100–101. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
