

Altered cohesin gene dosage affects Mammalian meiotic chromosome structure and behavior
- PMID: 23408896
- PMCID: PMC3567145
- DOI: 10.1371/journal.pgen.1003241
Altered cohesin gene dosage affects Mammalian meiotic chromosome structure and behavior
Abstract
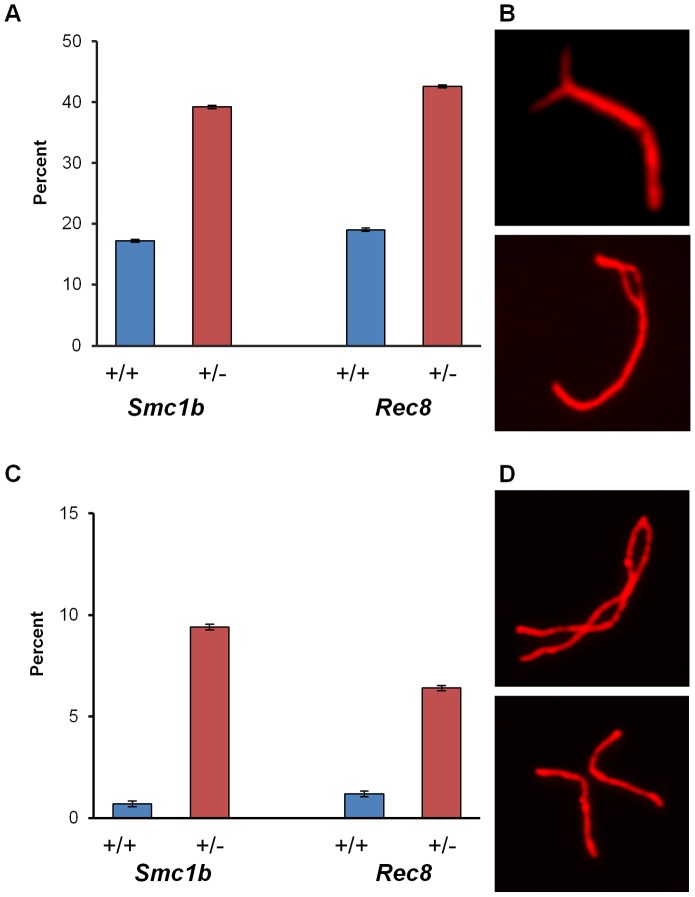
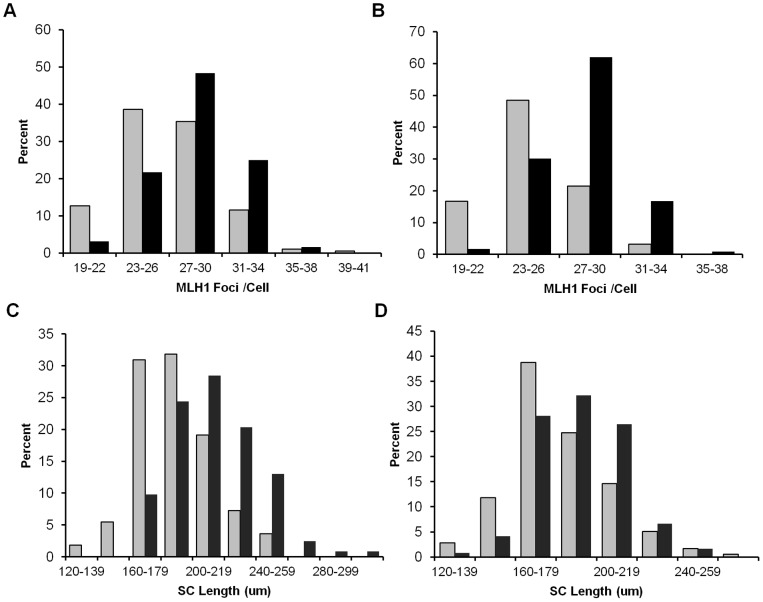
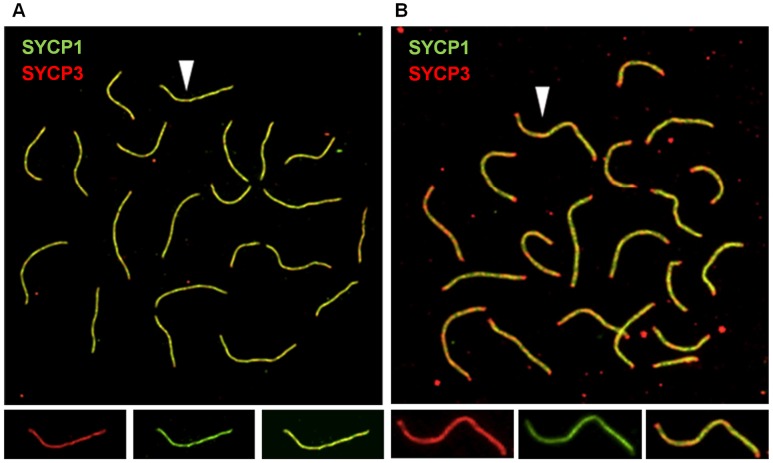
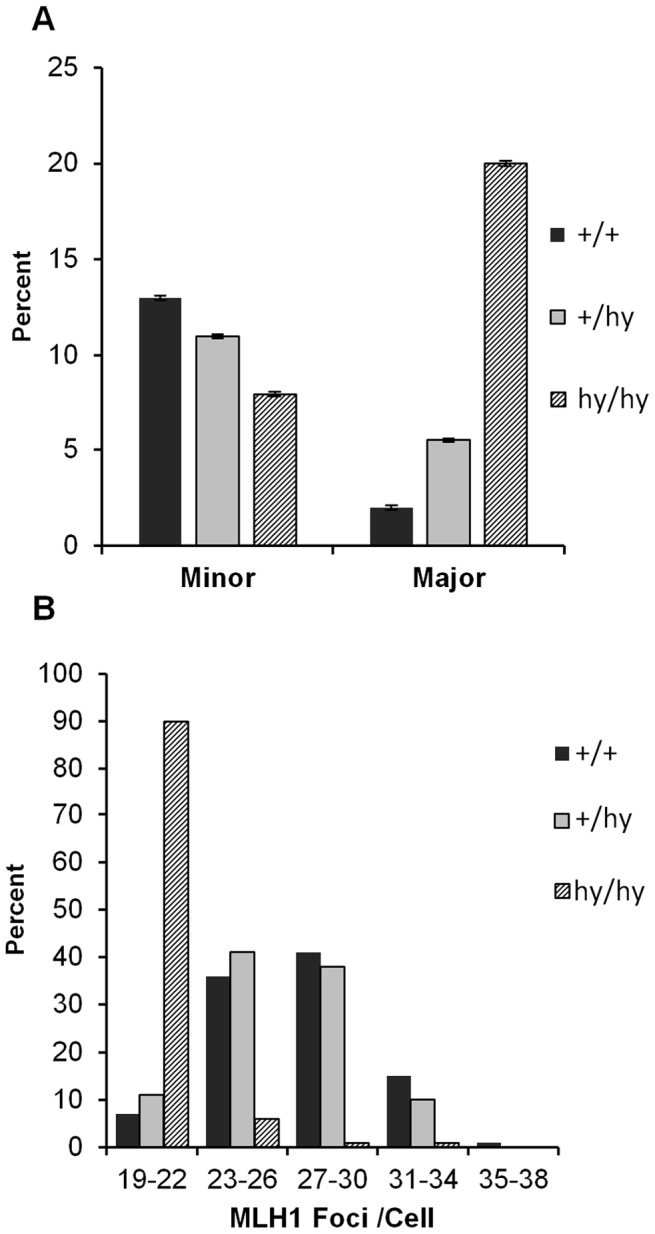
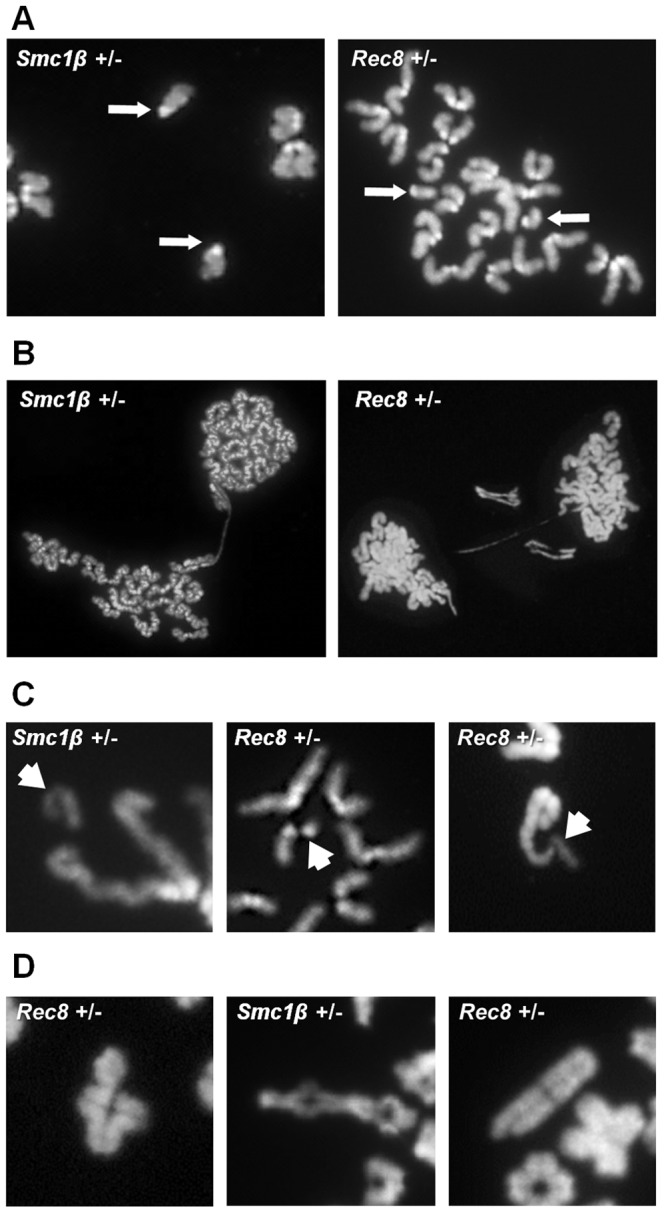
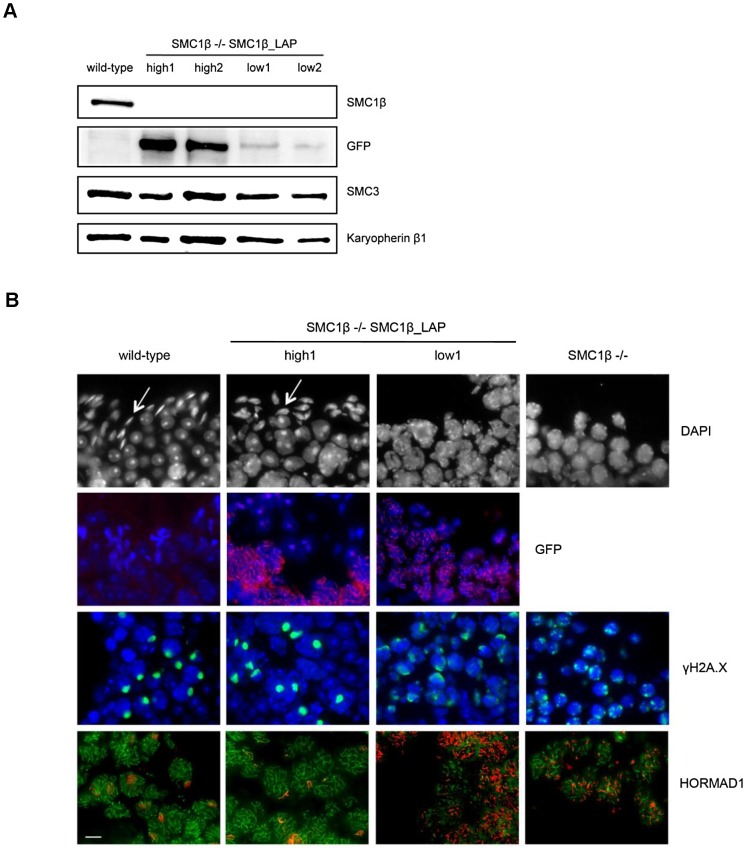
Based on studies in mice and humans, cohesin loss from chromosomes during the period of protracted meiotic arrest appears to play a major role in chromosome segregation errors during female meiosis. In mice, mutations in meiosis-specific cohesin genes cause meiotic disturbances and infertility. However, the more clinically relevant situation, heterozygosity for mutations in these genes, has not been evaluated. We report here evidence from the mouse that partial loss of gene function for either Smc1b or Rec8 causes perturbations in the formation of the synaptonemal complex (SC) and affects both synapsis and recombination between homologs during meiotic prophase. Importantly, these defects increase the frequency of chromosomally abnormal eggs in the adult female. These findings have important implications for humans: they suggest that women who carry mutations or variants that affect cohesin function have an elevated risk of aneuploid pregnancies and may even be at increased risk of transmitting structural chromosome abnormalities.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
STAG3-mediated stabilization of REC8 cohesin complexes promotes chromosome synapsis during meiosis.EMBO J. 2014 Jun 2;33(11):1243-55. doi: 10.1002/embj.201387329. Epub 2014 May 5. EMBO J. 2014. PMID: 24797475 Free PMC article.
-
Meiosis-specific cohesin component, Stag3 is essential for maintaining centromere chromatid cohesion, and required for DNA repair and synapsis between homologous chromosomes.PLoS Genet. 2014 Jul 3;10(7):e1004413. doi: 10.1371/journal.pgen.1004413. eCollection 2014 Jul. PLoS Genet. 2014. PMID: 24992337 Free PMC article.
-
Phosphorylation of chromosome core components may serve as axis marks for the status of chromosomal events during mammalian meiosis.PLoS Genet. 2012 Feb;8(2):e1002485. doi: 10.1371/journal.pgen.1002485. Epub 2012 Feb 9. PLoS Genet. 2012. PMID: 22346761 Free PMC article.
-
Meiotic prophase-like pathway for cleavage-independent removal of cohesin for chromosome morphogenesis.Curr Genet. 2019 Aug;65(4):817-827. doi: 10.1007/s00294-019-00959-x. Epub 2019 Mar 28. Curr Genet. 2019. PMID: 30923890 Review.
-
Rec8 Cohesin: A Structural Platform for Shaping the Meiotic Chromosomes.Genes (Basel). 2022 Jan 22;13(2):200. doi: 10.3390/genes13020200. Genes (Basel). 2022. PMID: 35205245 Free PMC article. Review.
Cited by
-
MicroRNA-202 safeguards meiotic progression by preventing premature SEPARASE-mediated REC8 cleavage.EMBO Rep. 2022 Aug 3;23(8):e54298. doi: 10.15252/embr.202154298. Epub 2022 Jun 17. EMBO Rep. 2022. PMID: 35712867 Free PMC article.
-
Localisation of the SMC loading complex Nipbl/Mau2 during mammalian meiotic prophase I.Chromosoma. 2014 Jun;123(3):239-52. doi: 10.1007/s00412-013-0444-7. Epub 2013 Nov 28. Chromosoma. 2014. PMID: 24287868 Free PMC article.
-
Meiotic Recombination: The Essence of Heredity.Cold Spring Harb Perspect Biol. 2015 Oct 28;7(12):a016618. doi: 10.1101/cshperspect.a016618. Cold Spring Harb Perspect Biol. 2015. PMID: 26511629 Free PMC article. Review.
-
SMC1α Substitutes for Many Meiotic Functions of SMC1β but Cannot Protect Telomeres from Damage.Curr Biol. 2018 Jan 22;28(2):249-261.e4. doi: 10.1016/j.cub.2017.12.020. Epub 2018 Jan 11. Curr Biol. 2018. PMID: 29337080 Free PMC article.
-
DNA damage-induced metaphase I arrest is mediated by the spindle assembly checkpoint and maternal age.Nat Commun. 2015 Nov 2;6:8706. doi: 10.1038/ncomms9706. Nat Commun. 2015. PMID: 26522734 Free PMC article.
References
-
- Hassold T, Hunt P (2001) To err (meiotically) is human: the genesis of human aneuploidy. Nat Rev Genet 2: 280–291. - PubMed
-
- Hassold T, Hall H, Hunt P (2007) The origin of human aneuploidy: where we have been, where we are going. Hum Mol Genet 16 Spec No. 2: R203–208. - PubMed
-
- Homer H (2007) Ageing aneuploidy and meiosis: eggs in a race against time.:. Yearbook of Obstetrics and Gynaecology 139–158.
-
- Hodges CA, Revenkova E, Jessberger R, Hassold TJ, Hunt PA (2005) SMC1beta-deficient female mice provide evidence that cohesins are a missing link in age-related nondisjunction. Nat Genet 37: 1351–1355. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
