

Inflammation-mediated genetic and epigenetic alterations drive cancer development in the neighboring epithelium upon stromal abrogation of TGF-β signaling
- PMID: 23408900
- PMCID: PMC3567148
- DOI: 10.1371/journal.pgen.1003251
Inflammation-mediated genetic and epigenetic alterations drive cancer development in the neighboring epithelium upon stromal abrogation of TGF-β signaling
Abstract
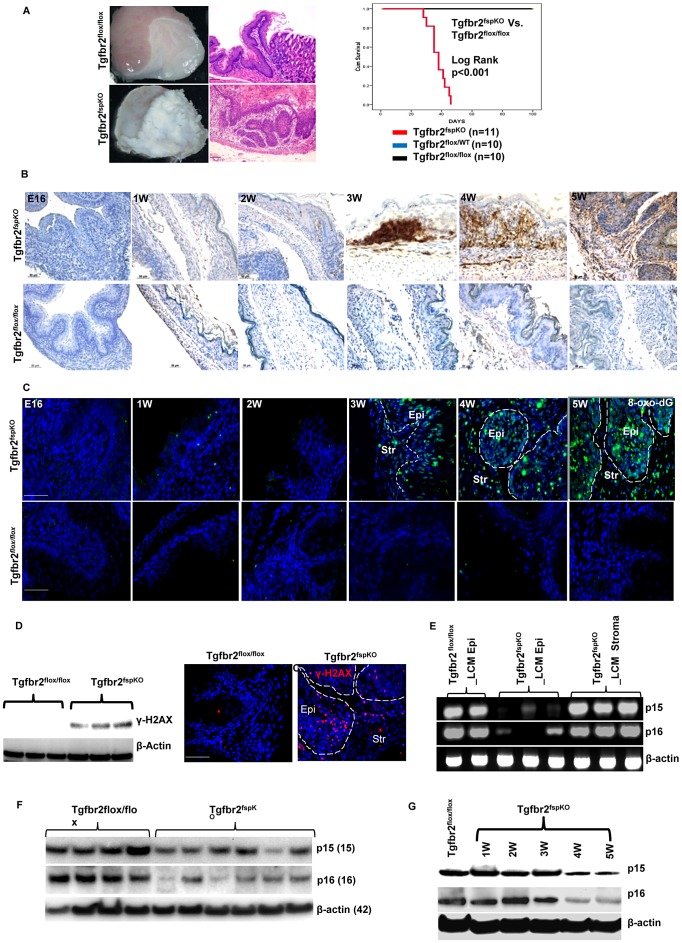
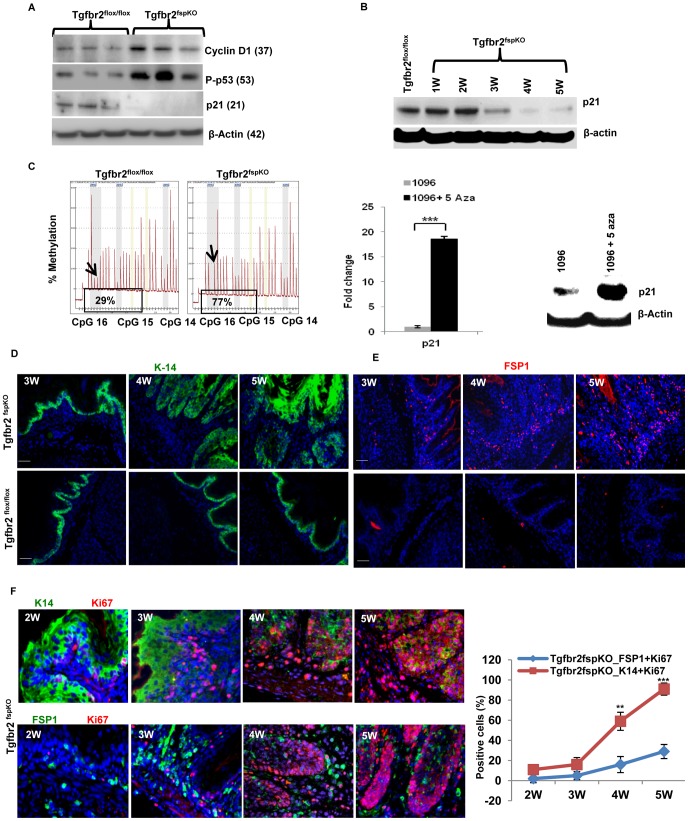
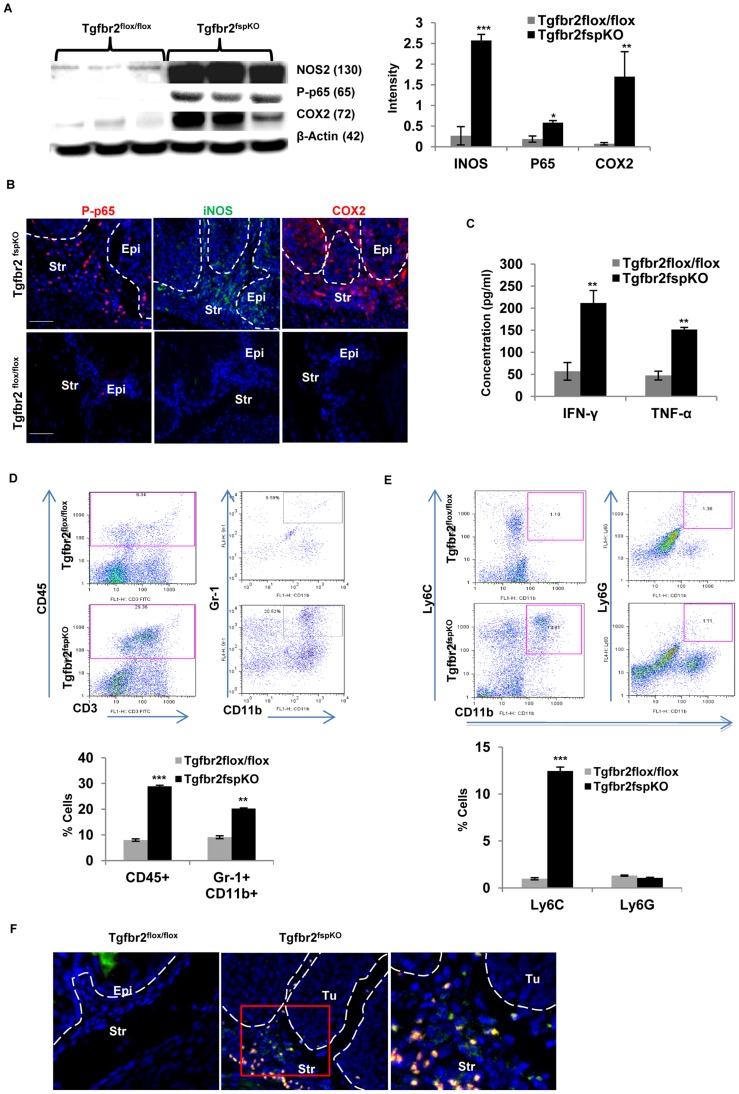
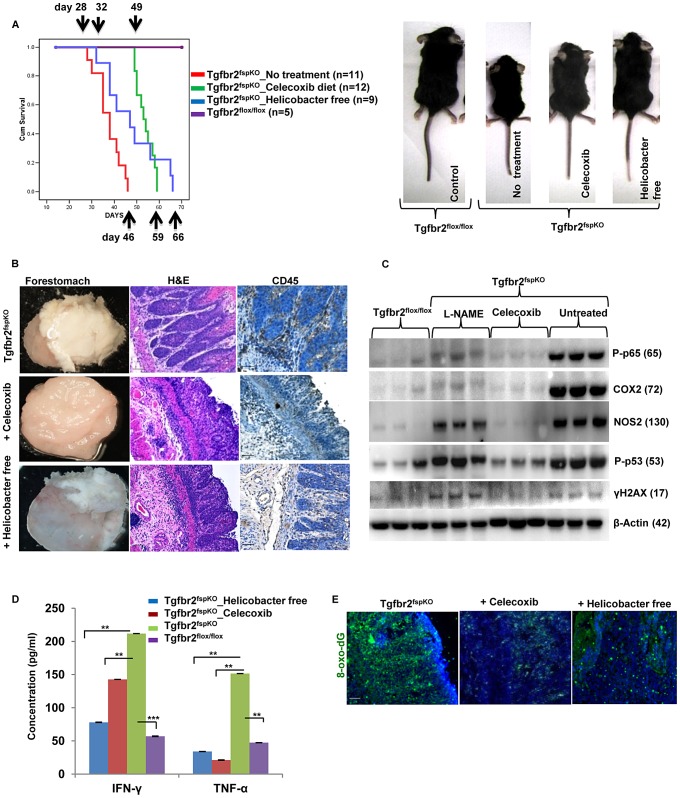
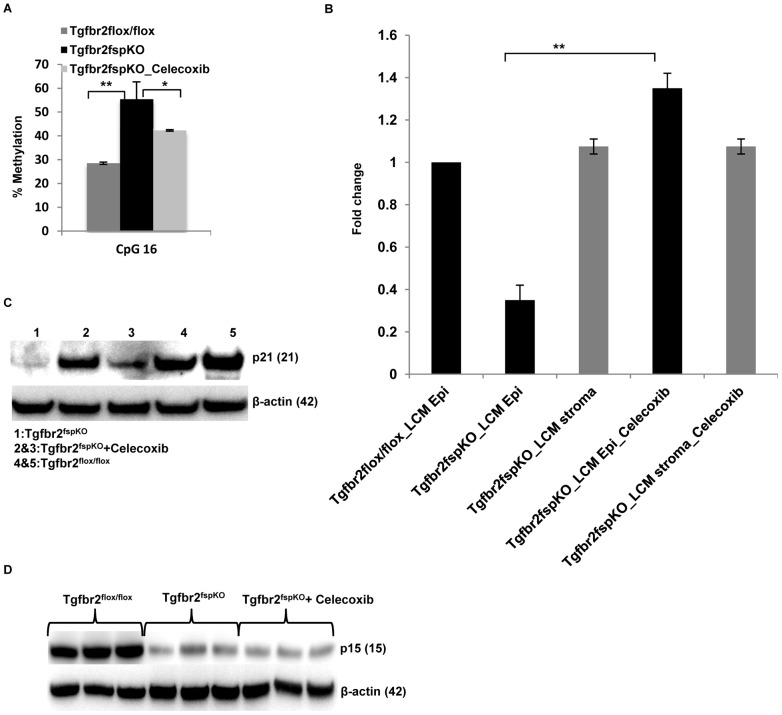
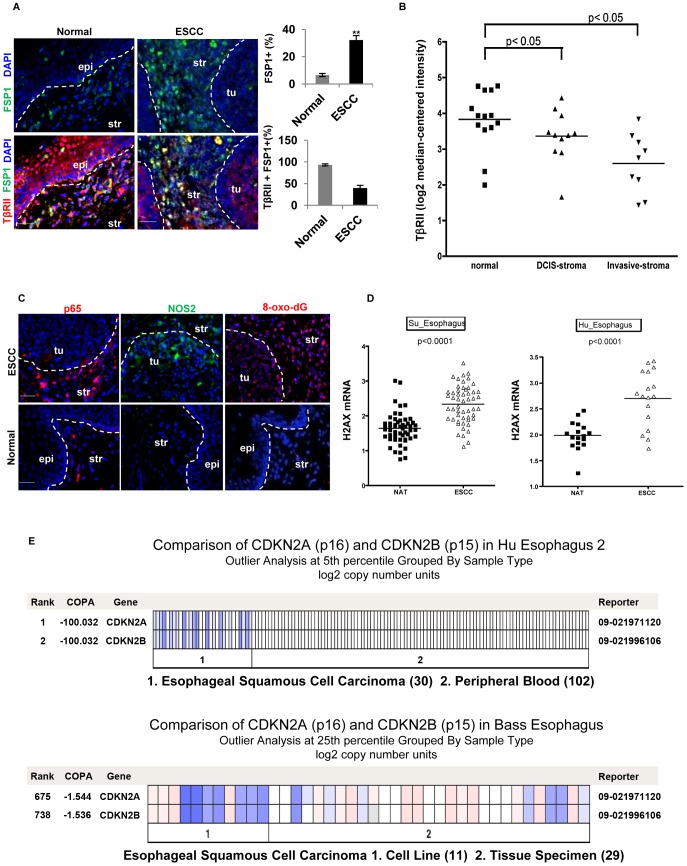
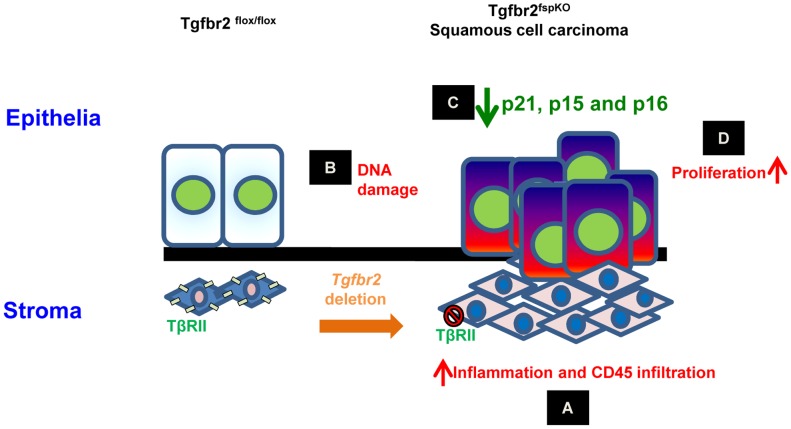
Deletion of tumor suppressor genes in stromal fibroblasts induces epithelial cancer development, suggesting an important role of stroma in epithelial homoeostasis. However, the underlying mechanisms remain to be elucidated. Here we report that deletion of the gene encoding TGFβ receptor 2 (Tgfbr2) in the stromal fibroblasts (Tgfbr2(fspKO)) induces inflammation and significant DNA damage in the neighboring epithelia of the forestomach. This results in loss or down-regulation of cyclin-dependent kinase inhibitors p15, p16, and p21, which contribute to the development of invasive squamous cell carcinoma (SCC). Anti-inflammation treatment restored p21 expression, delayed tumorigenesis, and increased survival of Tgfbr2(fspKO) mice. Our data demonstrate for the first time that inflammation is a critical player in the epigenetic silencing of p21 in tumor progression. Examination of human esophageal SCC showed a down-regulation of TGFβ receptor 2 (TβRII) in the stromal fibroblasts, as well as increased inflammation, DNA damage, and loss or decreased p15/p16 expression. Our study suggests anti-inflammation may be a new therapeutic option in treating human SCCs with down-regulation of TβRII in the stroma.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Comment in
-
Microenvironment: Making connections.Nat Rev Cancer. 2013 Apr;13(4):222-3. doi: 10.1038/nrc3492. Epub 2013 Mar 14. Nat Rev Cancer. 2013. PMID: 23486240 No abstract available.
References
-
- Bhowmick NA, Chytil A, Plieth D, Gorska AE, Dumont N, et al. (2004) TGF-beta signaling in fibroblasts modulates the oncogenic potential of adjacent epithelia. Science 303: 848–851. - PubMed
-
- Hill R, Song Y, Cardiff RD, Van Dyke T (2005) Selective evolution of stromal mesenchyme with p53 loss in response to epithelial tumorigenesis. Cell 123: 1001–1011. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
