

Evolution of predator dispersal in relation to spatio-temporal prey dynamics: how not to get stuck in the wrong place!
- PMID: 23408940
- PMCID: PMC3569443
- DOI: 10.1371/journal.pone.0054453
Evolution of predator dispersal in relation to spatio-temporal prey dynamics: how not to get stuck in the wrong place!
Abstract
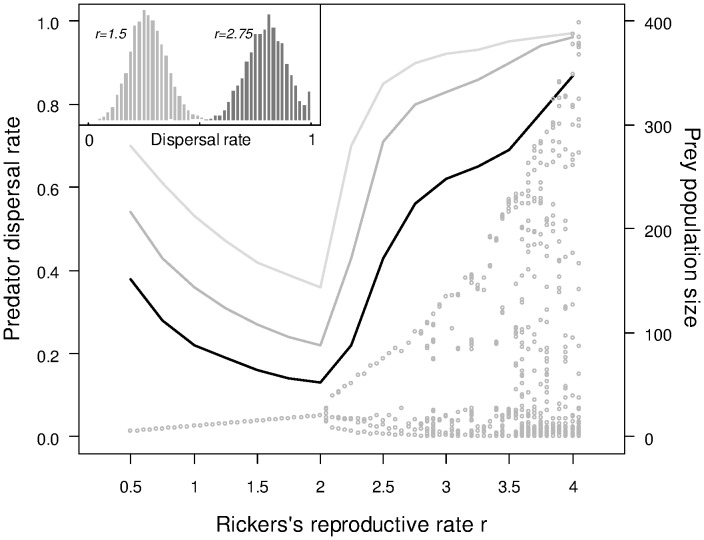
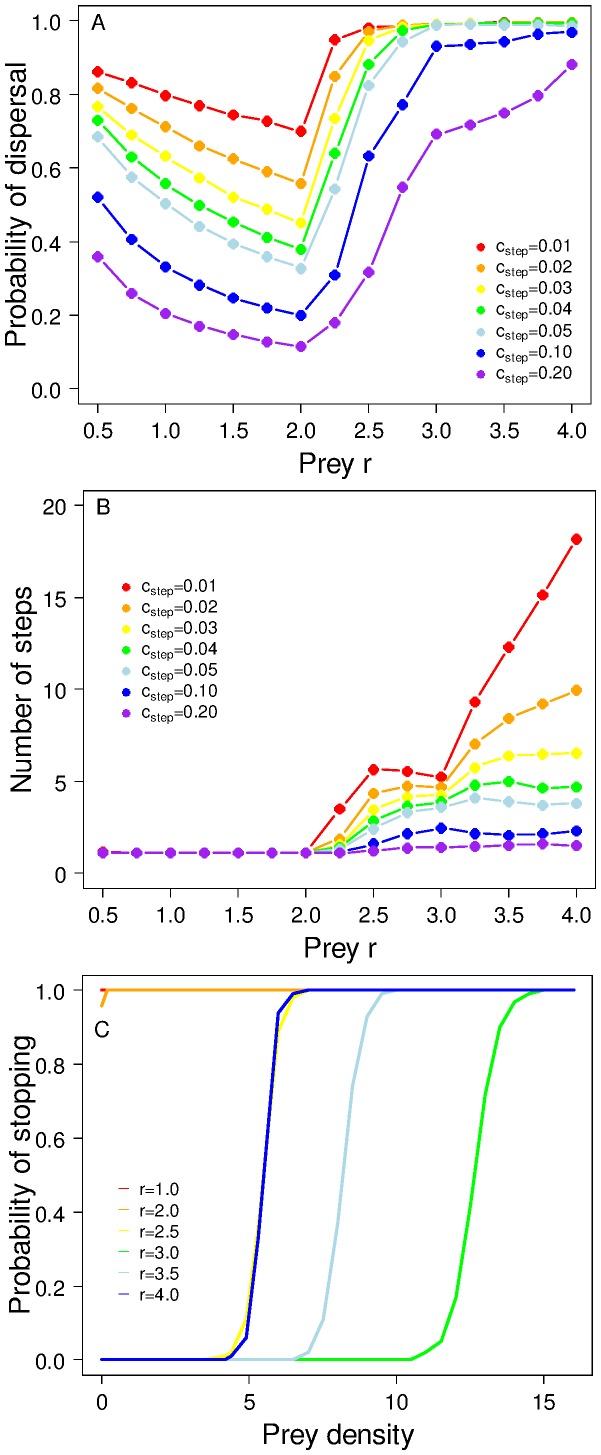
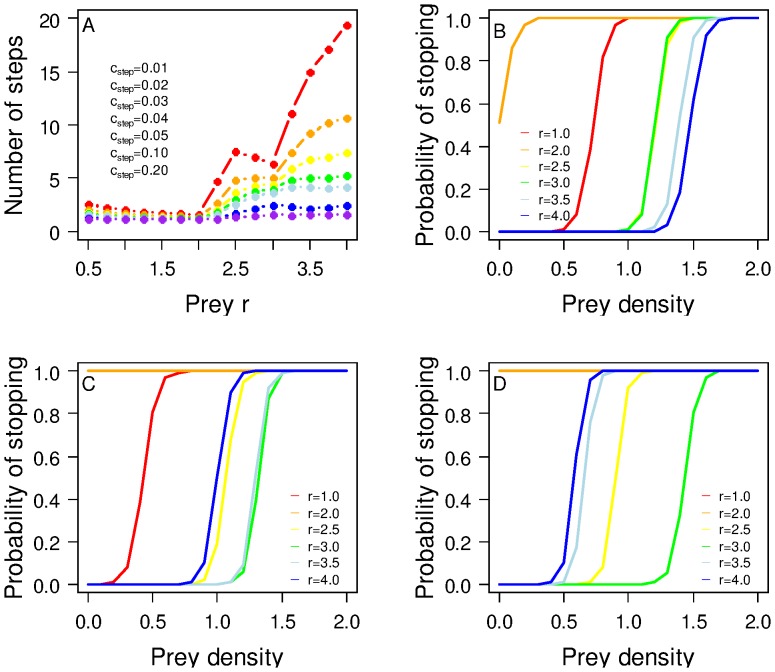
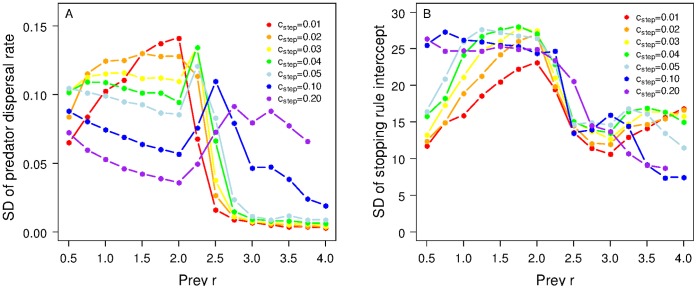
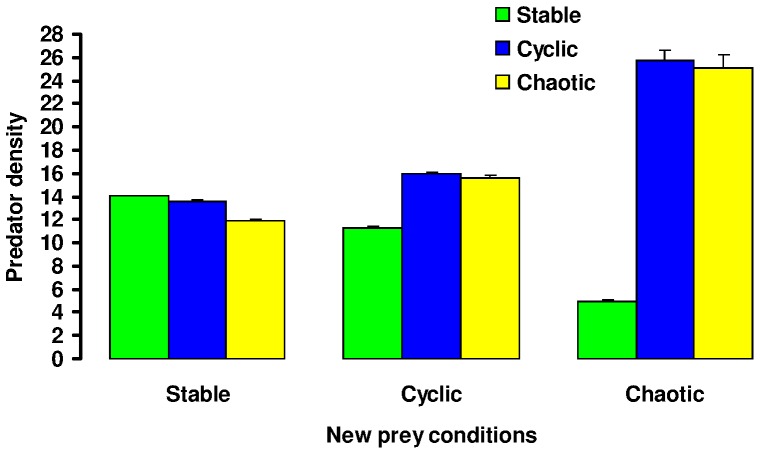
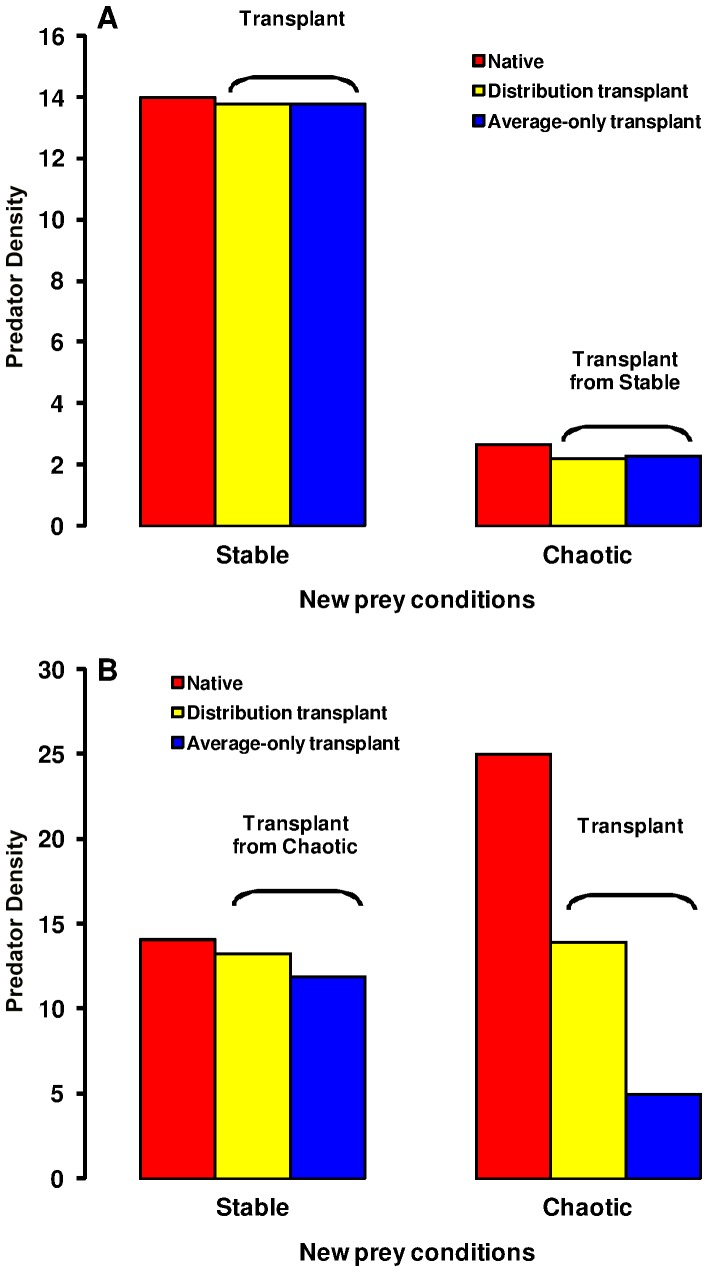
The eco-evolutionary dynamics of dispersal are recognised as key in determining the responses of populations to environmental changes. Here, by developing a novel modelling approach, we show that predators are likely to have evolved to emigrate more often and become more selective over their destination patch when their prey species exhibit spatio-temporally complex dynamics. We additionally demonstrate that the cost of dispersal can vary substantially across space and time. Perhaps as a consequence of current environmental change, many key prey species are currently exhibiting major shifts in their spatio-temporal dynamics. By exploring similar shifts in silico, we predict that predator populations will be most vulnerable when prey dynamics shift from stable to complex. The more sophisticated dispersal rules, and greater variance therein, that evolve under complex dynamics will enable persistence across a broader range of prey dynamics than the rules which evolve under relatively stable prey conditions.
Conflict of interest statement
Figures
References
-
- Bowler DE, Benton TG (2005) Causes and consequences of animal dispersal strategies: relating individual behaviour to spatial dynamics. Biol. Rev. Cambridge Phil. Soc. 80: 205–225. - PubMed
-
- Simmons AD, Thomas CD (2004) Changes in dispersal during species’ range expansions. Am. Nat. 164: 378–395. - PubMed
-
- Phillips BL, Brown GP, Webb JK, Shine R (2006) Invasion and the evolution of speed in toads. Nature 439: 803. - PubMed
-
- Lachmuth S, Durka W, Schurr FM (2011) Differentiation of reproductive and competitive ability in the invaded range of Senecio inaequidens: the role of genetic Allee effects, adaptive and nonadaptive evolution. New Phyt 192: 529–541 doi:10.1111/j.1469–8137.2011.03808.x. - DOI - PubMed
-
- Kokko H, Lopez-Sepulcre A (2006) From Individual Dispersal to Species Ranges: Perspectives for a Changing World. Science 313: 789–791 doi:10.1126/science.1128566. - DOI - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
