

Differential roles of Hath1, MUC2 and P27Kip1 in relation with gamma-secretase inhibition in human colonic carcinomas: a translational study
- PMID: 23409082
- PMCID: PMC3569436
- DOI: 10.1371/journal.pone.0055904
Differential roles of Hath1, MUC2 and P27Kip1 in relation with gamma-secretase inhibition in human colonic carcinomas: a translational study
Abstract
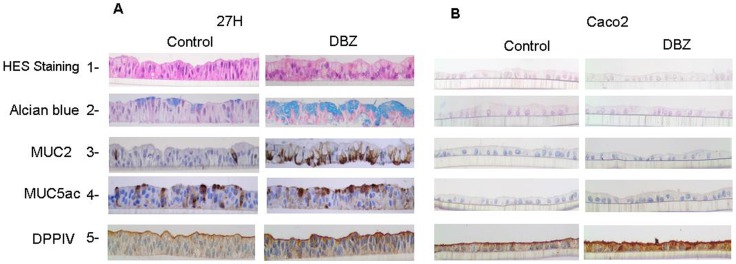
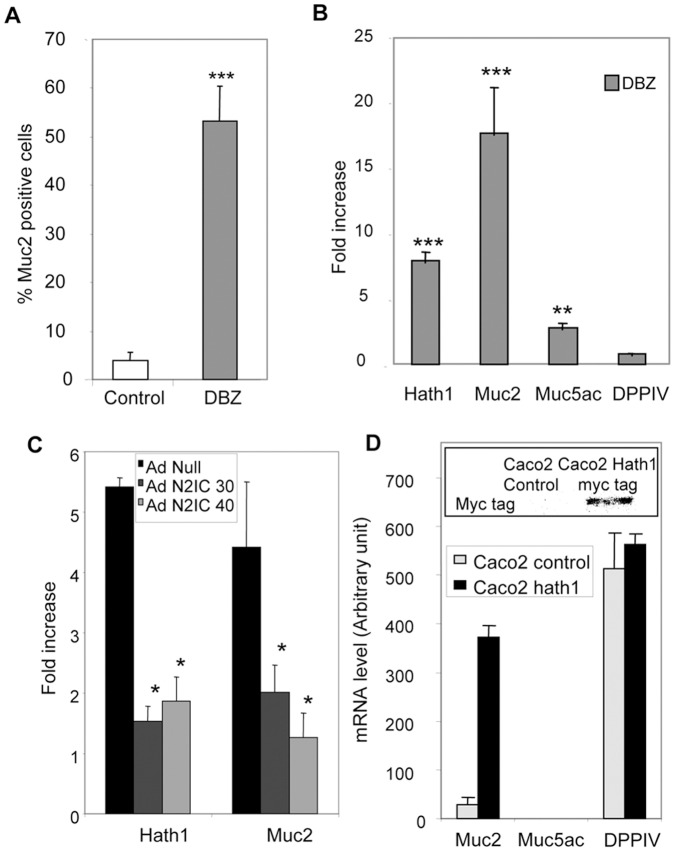
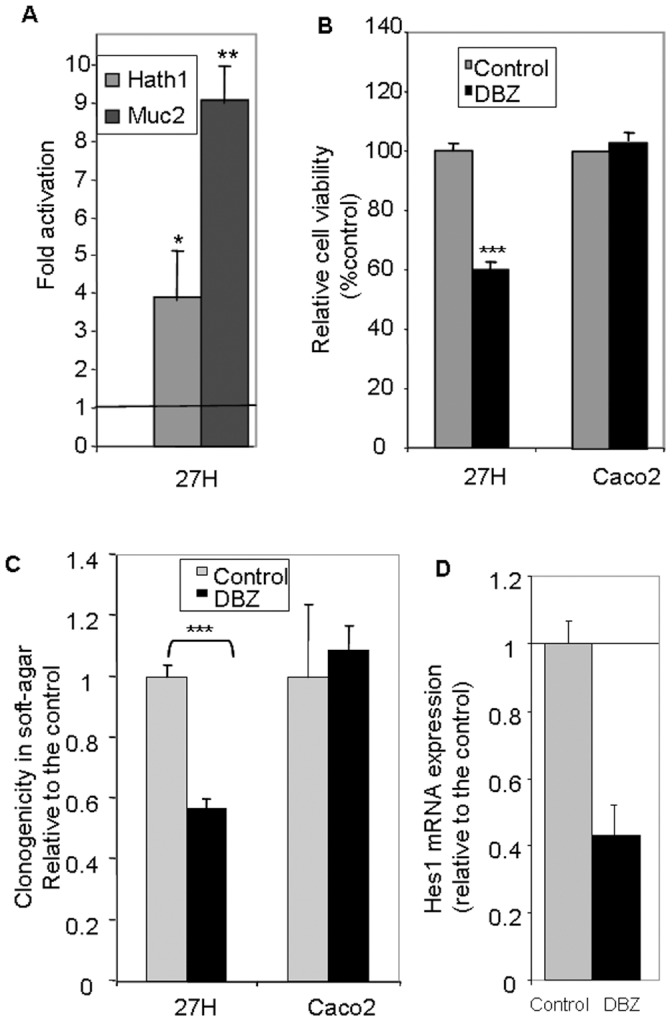
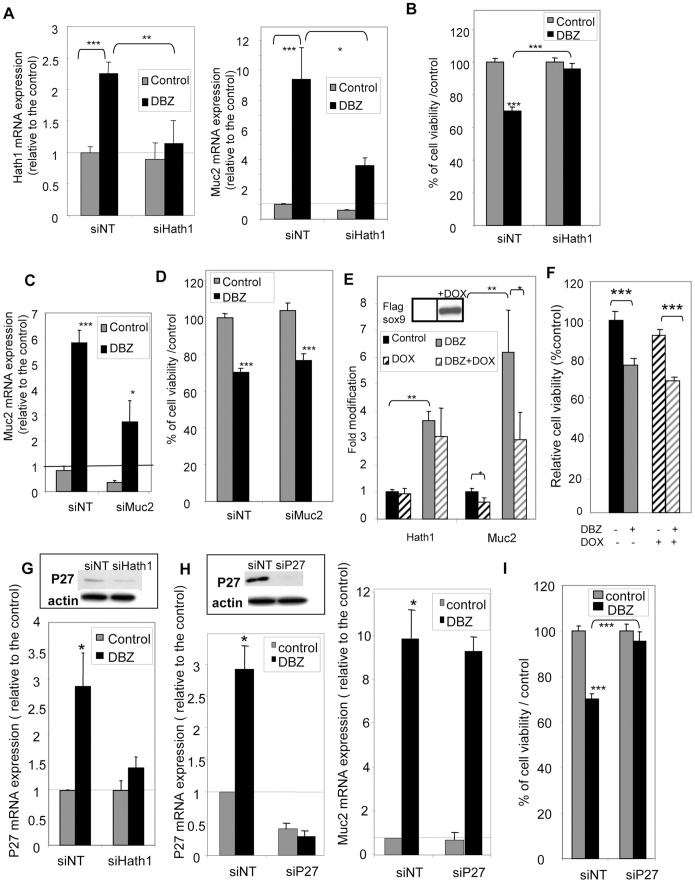
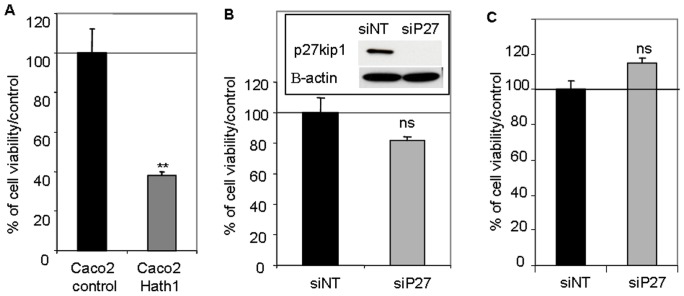
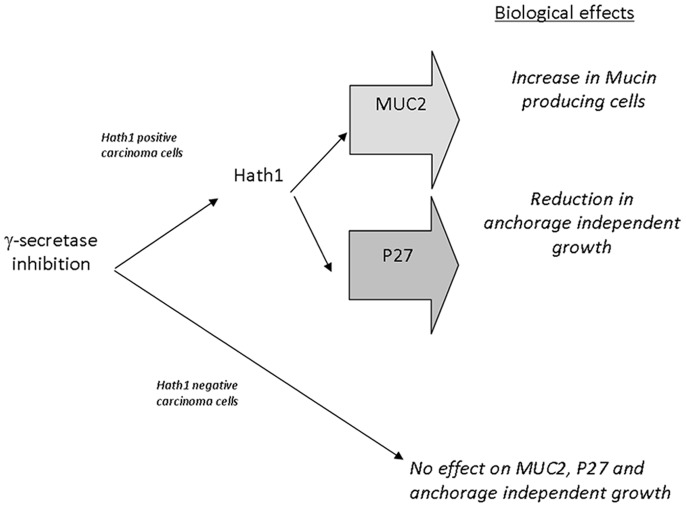
Hath1, a bHLH transcription factor negatively regulated by the γ-secretase-dependent Notch pathway, is required for intestinal secretory cell differentiation. Our aim was fourfold: 1) determine whether Hath1 is able to alter the phenotype of colon cancer cells that are committed to a differentiated phenotype, 2) determine whether the Hath1-dependent alteration of differentiation is coupled to a restriction of anchorage-dependent growth, 3) decipher the respective roles of three putative tumor suppressor genes Hath1, MUC2 and P27kip1 in this coupling and, 4) examine how our findings translate to primary tumors. Human colon carcinoma cell lines that differentiate along a mucin secreting (MUC2/MUC5AC) and/or enterocytic (DPPIV) lineages were maintained on inserts with or without a γ-secretase inhibitor (DBZ). Then the cells were detached and their ability to survive/proliferate in the absence of substratum was assessed. γ-secretase inhibition led to a Hath1-mediated preferential induction of MUC2 over MUC5AC, without DPPIV modification, in association with a decrease in anchorage-independent growth. While P27kip1 silencing relieved the cells from the Hath1-induced decrease of anchorage-independent growth, MUC2 silencing did not modify this parameter. Hath1 ectopic expression in the Hath1 negative enterocytic Caco2 cells led to a decreased anchorage-independent growth in a P27kip1-independent manner. In cultured primary human colon carcinomas, Hath1 was up-regulated in 7 out of 10 tumors upon DBZ treatment. Parallel MUC2 up-regulation occurred in 4 (4/7) and P27kip1 in only 2 (2/7) tumors. Interestingly, the response patterns of primary tumors to DBZ fitted with the hierarchical model of divergent signalling derived from our findings on cell lines.
Conflict of interest statement
Figures
Similar articles
-
Hath1 inhibits proliferation of colon cancer cells probably through up-regulating expression of Muc2 and p27 and down-regulating expression of cyclin D1.Asian Pac J Cancer Prev. 2012;13(12):6349-55. doi: 10.7314/apjcp.2012.13.12.6349. Asian Pac J Cancer Prev. 2012. PMID: 23464457
-
Hath1, down-regulated in colon adenocarcinomas, inhibits proliferation and tumorigenesis of colon cancer cells.Cancer Res. 2004 Sep 1;64(17):6050-7. doi: 10.1158/0008-5472.CAN-04-0290. Cancer Res. 2004. PMID: 15342386
-
A role for Hath1, a bHLH transcription factor, in colon adenocarcinoma.Ann N Y Acad Sci. 2005 Nov;1059:174-83. doi: 10.1196/annals.1339.048. Ann N Y Acad Sci. 2005. PMID: 16382053 Review.
-
Suppression of hath1 gene expression directly regulated by hes1 via notch signaling is associated with goblet cell depletion in ulcerative colitis.Inflamm Bowel Dis. 2011 Nov;17(11):2251-60. doi: 10.1002/ibd.21611. Epub 2011 Jan 6. Inflamm Bowel Dis. 2011. PMID: 21987298
-
Bacteria regulate intestinal epithelial cell differentiation factors both in vitro and in vivo.PLoS One. 2013;8(2):e55620. doi: 10.1371/journal.pone.0055620. Epub 2013 Feb 13. PLoS One. 2013. PMID: 23418447 Free PMC article.
Cited by
-
MUC4 is negatively regulated through the Wnt/β-catenin pathway via the Notch effector Hath1 in colorectal cancer.Genes Cancer. 2016 May;7(5-6):154-168. doi: 10.18632/genesandcancer.108. Genes Cancer. 2016. PMID: 27551331 Free PMC article.
-
Mechanistic and Functional Shades of Mucins and Associated Glycans in Colon Cancer.Cancers (Basel). 2020 Mar 11;12(3):649. doi: 10.3390/cancers12030649. Cancers (Basel). 2020. PMID: 32168759 Free PMC article. Review.
-
Subversion of human intestinal mucosa innate immunity by a Crohn's disease-associated E. coli.Mucosal Immunol. 2015 May;8(3):572-81. doi: 10.1038/mi.2014.89. Epub 2014 Oct 1. Mucosal Immunol. 2015. PMID: 25269707
-
Atonal bHLH transcription factor 1 is an important factor for maintaining the balance of cell proliferation and differentiation in tumorigenesis.Oncol Lett. 2020 Sep;20(3):2595-2605. doi: 10.3892/ol.2020.11833. Epub 2020 Jul 8. Oncol Lett. 2020. PMID: 32782577 Free PMC article. Review.
References
-
- De Strooper B, Annaert W, Cupers P, Saftig P, Craessaerts K, et al. (1999) A presenilin-1-dependent gamma-secretase-like protease mediates release of Notch intracellular domain. Nature 398: 518–522. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
