

A cis-acting element in retroviral genomic RNA links Gag-Pol ribosomal frameshifting to selective viral RNA encapsidation
- PMID: 23414758
- PMCID: PMC3587049
- DOI: 10.1016/j.chom.2013.01.007
A cis-acting element in retroviral genomic RNA links Gag-Pol ribosomal frameshifting to selective viral RNA encapsidation
Abstract
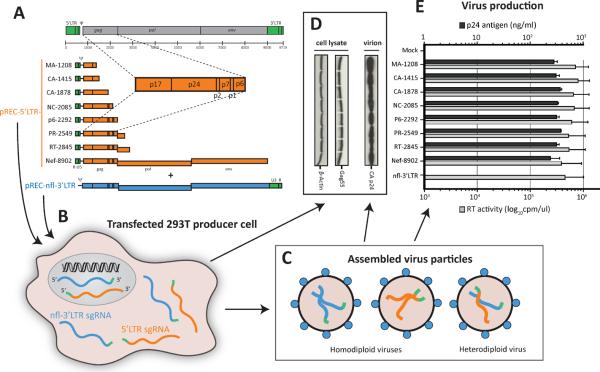
During retroviral RNA encapsidation, two full-length genomic (g) RNAs are selectively incorporated into assembling virions. Packaging involves a cis-acting packaging element (Ψ) within the 5' untranslated region of unspliced HIV-1 RNA genome. However, the mechanism(s) that selects and limits gRNAs for packaging remains uncertain. Using a dual complementation system involving bipartite HIV-1 gRNA, we observed that gRNA packaging is additionally dependent on a cis-acting RNA element, the genomic RNA packaging enhancer (GRPE), found within the gag p1-p6 domain and overlapping the Gag-Pol ribosomal frameshift signal. Deleting or disrupting the two conserved GRPE stem loops diminished gRNA packaging and infectivity >50-fold, while deleting gag sequences between Ψ and GRPE had no effect. Downregulating the translation termination factor eRF1 produces defective virus particles containing 20 times more gRNA. Thus, only the HIV-1 RNAs employed for Gag-Pol translation may be specifically selected for encapsidation, possibly explaining the limitation of two gRNAs per virion.
Copyright © 2013 Elsevier Inc. All rights reserved.
Figures






Comment in
-
HIV-1: packaging a shifty genome?Cell Host Microbe. 2013 Feb 13;13(2):123-5. doi: 10.1016/j.chom.2013.01.015. Cell Host Microbe. 2013. PMID: 23414752
References
-
- Abbink TE, Berkhout B. A novel long distance base-pairing interaction in human immunodeficiency virus type 1 RNA occludes the Gag start codon. J. Biol. Chem. 2003;278:11601–11611. - PubMed
-
- Amarasinghe GK, De Guzman RN, Turner RB, Chancellor KJ, Wu ZR, Summers MF. NMR structure of the HIV-1 nucleocapsid protein bound to stem-loop SL2 of the psi-RNA packaging signal. Implications for genome recognition. J. Mol. Biol. 2000;301:491–511. - PubMed
-
- Berkowitz R, Fisher J, Goff SP. RNA packaging. Curr. Top. Microbiol. Immunol. 1996;214:177–218. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
