

EGF transactivation of Trk receptors regulates the migration of newborn cortical neurons
- PMID: 23416450
- PMCID: PMC4148818
- DOI: 10.1038/nn.3333
EGF transactivation of Trk receptors regulates the migration of newborn cortical neurons
Abstract
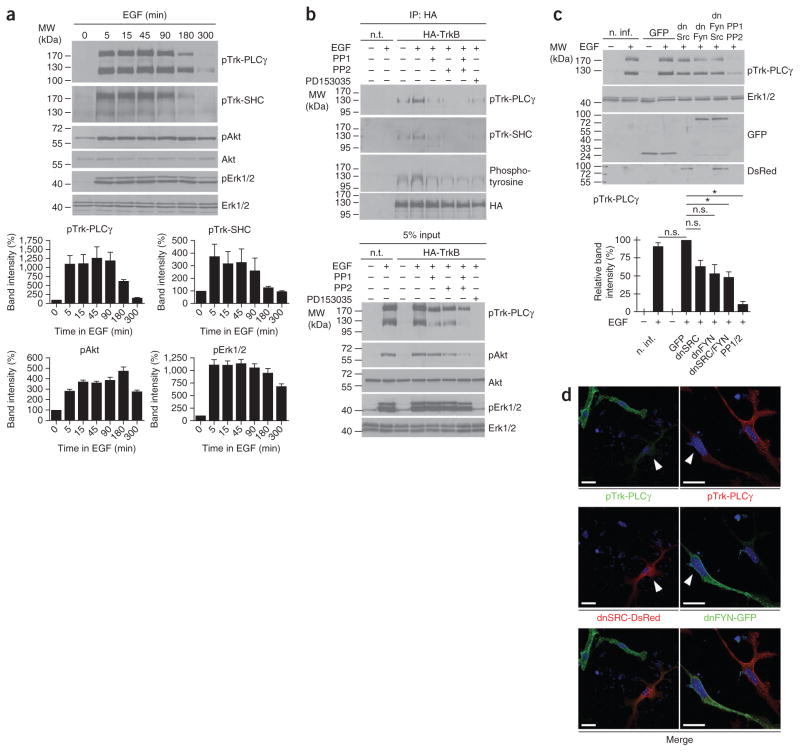
The development of neuronal networks in the neocortex depends on control mechanisms for mitosis and migration that allow newborn neurons to find their accurate position. Multiple mitogens, neurotrophic factors, guidance molecules and their corresponding receptors are involved in this process, but the mechanisms by which these signals are integrated are only poorly understood. We found that TrkB and TrkC, the receptors for brain-derived neurotrophic factor (BDNF) and neurotrophin-3 (NT-3), are activated by epidermal growth factor receptor (EGFR) signaling rather than by BDNF or NT-3 in embryonic mouse cortical precursor cells. This transactivation event regulated migration of early neuronal cells to their final position in the developing cortex. Transactivation by EGF led to membrane translocation of TrkB, promoting its signaling responsiveness. Our results provide genetic evidence that TrkB and TrkC activation in early cortical neurons do not depend on BDNF and NT-3, but instead on transactivation by EGFR signaling.
Conflict of interest statement
The authors declare no competing financial interests.
Figures





References
-
- Doetsch F, Caille I, Lim DA, Garcia-Verdugo JM, Alvarez-Buylla A. Subventricular zone astrocytes are neural stem cells in the adult mammalian brain. Cell. 1999;97:703–716. - PubMed
-
- Götz M, Huttner WB. The cell biology of neurogenesis. Nat Rev Mol Cell Biol. 2005;6:777–788. - PubMed
-
- Alvarez-Buylla A, Garcia-Verdugo JM, Tramontin AD. A unified hypothesis on the lineage of neural stem cells. Nat Rev Neurosci. 2001;2:287–293. - PubMed
-
- Gupta A, Tsai LH, Wynshaw-Boris A. Life is a journey: a genetic look at neocortical development. Nat Rev Genet. 2002;3:342–355. - PubMed
-
- Angevine JB, Sidman RL. Autoradiographic study of cell migration during histogenesis of cerebral cortex in the mouse. Nature. 1961;192:766–768. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
