

Crystal structure of the entire respiratory complex I
- PMID: 23417064
- PMCID: PMC3672946
- DOI: 10.1038/nature11871
Crystal structure of the entire respiratory complex I
Abstract
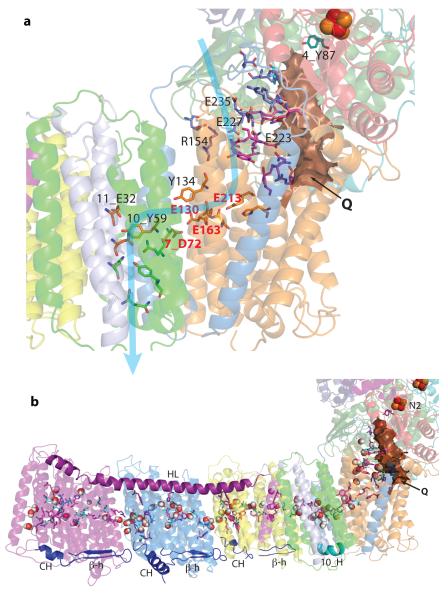
Complex I is the first and largest enzyme of the respiratory chain and has a central role in cellular energy production through the coupling of NADH:ubiquinone electron transfer to proton translocation. It is also implicated in many common human neurodegenerative diseases. Here, we report the first crystal structure of the entire, intact complex I (from Thermus thermophilus) at 3.3 Å resolution. The structure of the 536-kDa complex comprises 16 different subunits, with a total of 64 transmembrane helices and 9 iron-sulphur clusters. The core fold of subunit Nqo8 (ND1 in humans) is, unexpectedly, similar to a half-channel of the antiporter-like subunits. Small subunits nearby form a linked second half-channel, which completes the fourth proton-translocation pathway (present in addition to the channels in three antiporter-like subunits). The quinone-binding site is unusually long, narrow and enclosed. The quinone headgroup binds at the deep end of this chamber, near iron-sulphur cluster N2. Notably, the chamber is linked to the fourth channel by a 'funnel' of charged residues. The link continues over the entire membrane domain as a flexible central axis of charged and polar residues, and probably has a leading role in the propagation of conformational changes, aided by coupling elements. The structure suggests that a unique, out-of-the-membrane quinone-reaction chamber enables the redox energy to drive concerted long-range conformational changes in the four antiporter-like domains, resulting in translocation of four protons per cycle.
Figures




References
-
- Walker JE. The NADH - ubiquinone oxidoreductase (complex I) of respiratory chains. Q. Rev. Biophys. 1992;25:253–324. - PubMed
-
- Yagi T, Matsuno-Yagi A. The proton-translocating NADH-Quinone oxidoreductase in the respiratory chain: the secret unlocked. Biochemistry. 2003;42:2266–2274. - PubMed
-
- Brandt U. Energy converting NADH:quinone oxidoreductase (complex I) Annu. Rev. Biochem. 2006;75:69–92. - PubMed
-
- Ohnishi T. Iron-sulfur clusters/semiquinones in complex I. Biochim. Biophys. Acta. 1998;1364:186–206. - PubMed
-
- Sazanov LA. Respiratory complex I: mechanistic and structural insights provided by the crystal structure of the hydrophilic domain. Biochemistry. 2007;46:2275–2288. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
