

Isolation and functional characterization of human erythroblasts at distinct stages: implications for understanding of normal and disordered erythropoiesis in vivo
- PMID: 23422750
- PMCID: PMC3630836
- DOI: 10.1182/blood-2013-01-476390
Isolation and functional characterization of human erythroblasts at distinct stages: implications for understanding of normal and disordered erythropoiesis in vivo
Abstract
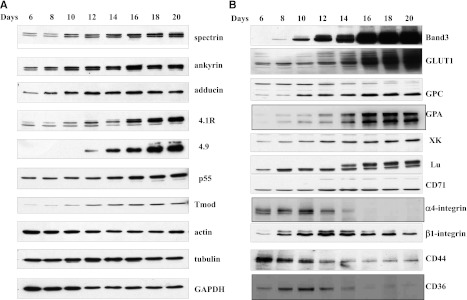
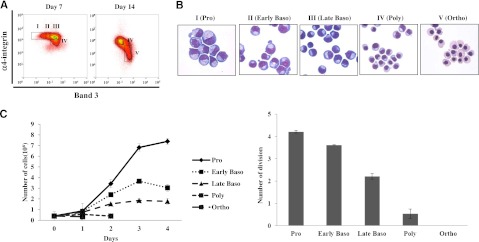
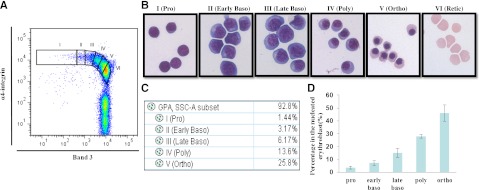
Terminal erythroid differentiation starts from morphologically recognizable proerythroblasts that proliferate and differentiate to generate red cells. Although this process has been extensively studied in mice, its characterization in humans is limited. By examining the dynamic changes of expression of membrane proteins during in vitro human terminal erythroid differentiation, we identified band 3 and α4 integrin as optimal surface markers for isolating 5 morphologically distinct populations at successive developmental stages. Functional analysis revealed that these purified cell populations have distinct mitotic capacity. Use of band 3 and α4 integrin enabled us to isolate erythroblasts at specific developmental stages from primary human bone marrow. The ratio of erythroblasts at successive stages followed the predicted 1:2:4:8:16 pattern. In contrast, bone marrows from myelodysplastic syndrome patients exhibited altered terminal erythroid differentiation profiles. Thus, our findings not only provide new insights into the genesis of the red cell membrane during human terminal erythroid differentiation but also offer a means of isolating and quantifying each developmental stage during terminal erythropoiesis in vivo. Our findings should facilitate a comprehensive cellular and molecular characterization of each specific developmental stage of human erythroblasts and should provide a powerful means of identifying stage-specific defects in diseases associated with pathological erythropoiesis.
Figures



References
-
- Blikstad I, Nelson WJ, Moon RT, Lazarides E. Synthesis and assembly of spectrin during avian erythropoiesis: stoichiometric assembly but unequal synthesis of alpha and beta spectrin. Cell. 1983;32(4):1081–1091. - PubMed
-
- Hanspal M, Hanspal JS, Kalraiya R, Liu SC, Sahr KE, Howard D, Palek J. Asynchronous synthesis of membrane skeletal proteins during terminal maturation of murine erythroblasts. Blood. 1992;80(2):530–539. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
