

Reciprocal regulation of protein synthesis and carbon metabolism for thylakoid membrane biogenesis
- PMID: 23424285
- PMCID: PMC3570535
- DOI: 10.1371/journal.pbio.1001482
Reciprocal regulation of protein synthesis and carbon metabolism for thylakoid membrane biogenesis
Abstract
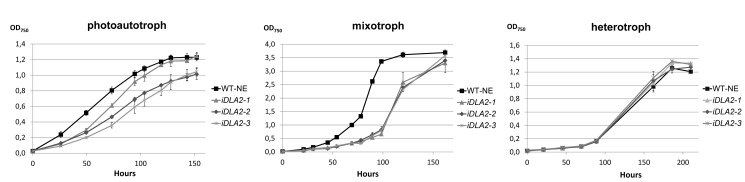
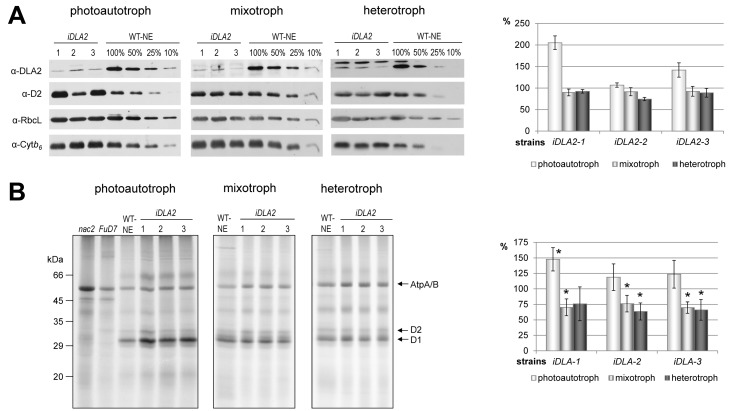
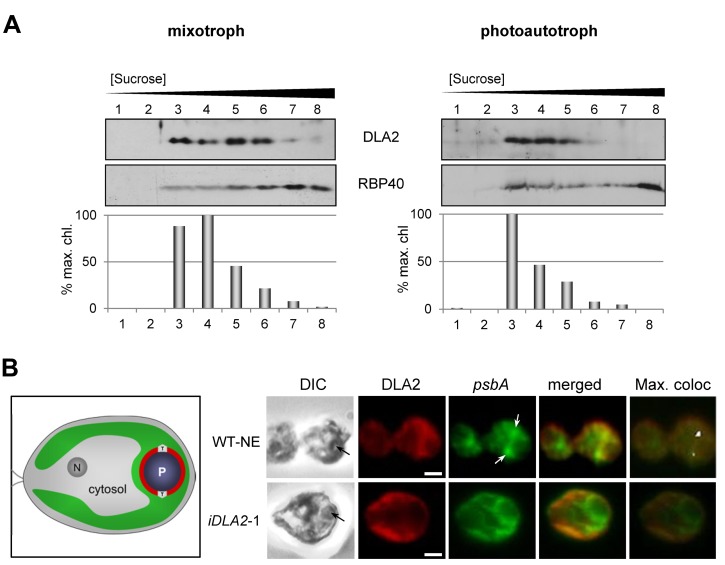
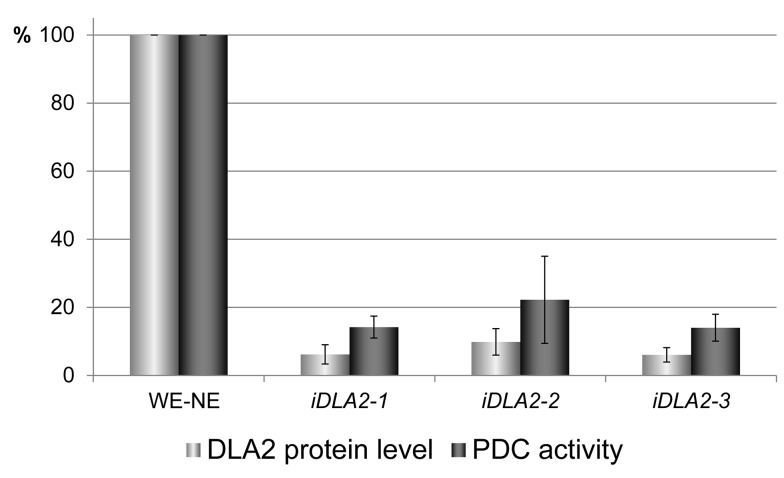
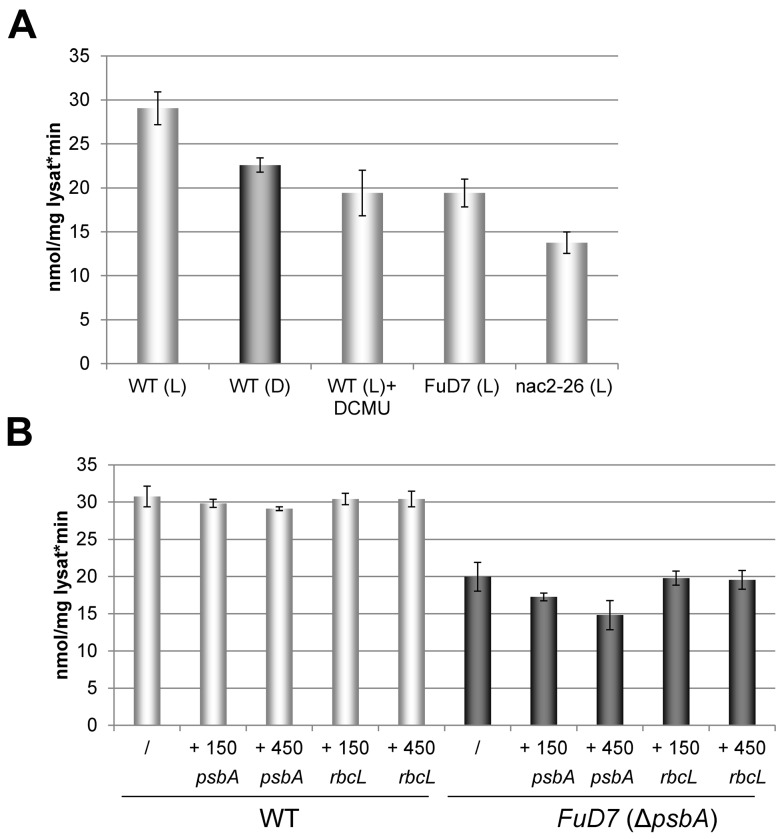
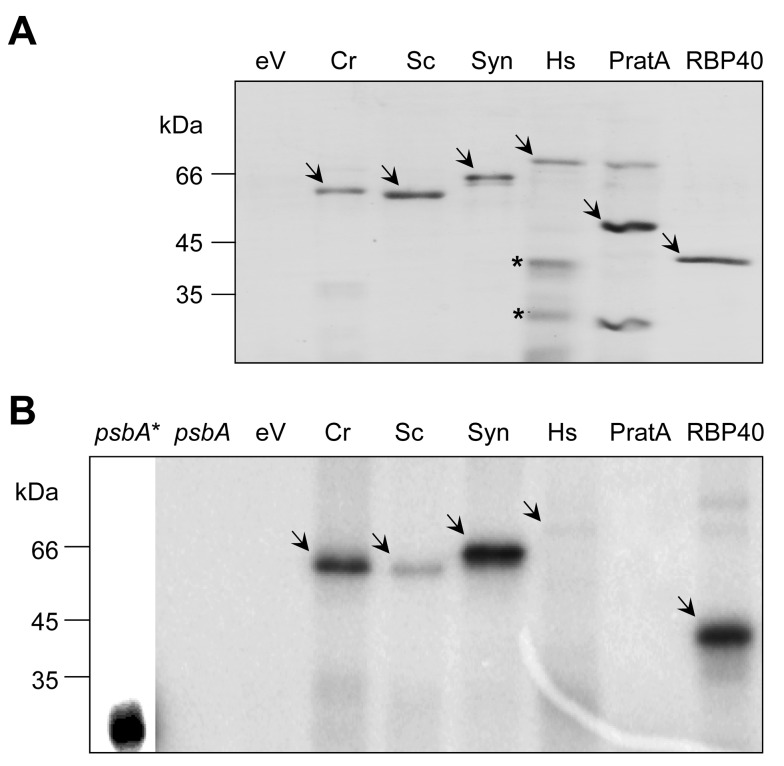
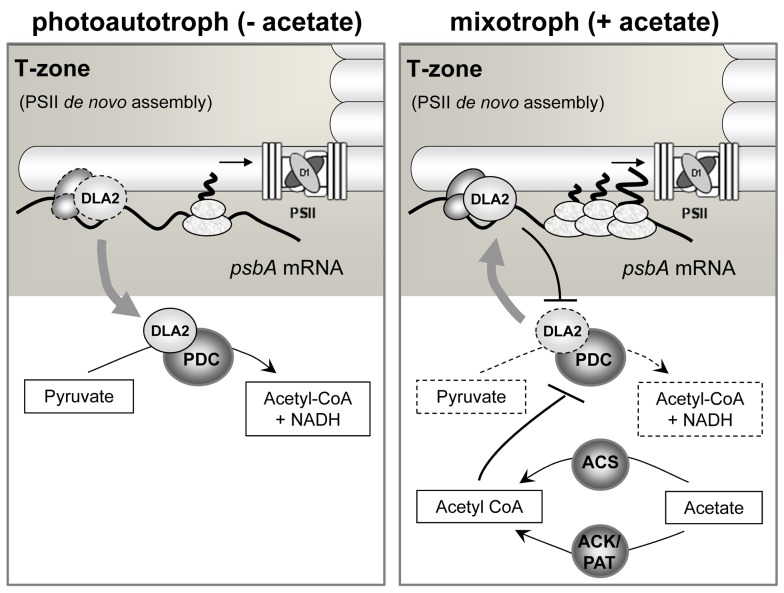
Metabolic control of gene expression coordinates the levels of specific gene products to meet cellular demand for their activities. This control can be exerted by metabolites acting as regulatory signals and/or a class of metabolic enzymes with dual functions as regulators of gene expression. However, little is known about how metabolic signals affect the balance between enzymatic and regulatory roles of these dual functional proteins. We previously described the RNA binding activity of a 63 kDa chloroplast protein from Chlamydomonas reinhardtii, which has been implicated in expression of the psbA mRNA, encoding the D1 protein of photosystem II. Here, we identify this factor as dihydrolipoamide acetyltransferase (DLA2), a subunit of the chloroplast pyruvate dehydrogenase complex (cpPDC), which is known to provide acetyl-CoA for fatty acid synthesis. Analyses of RNAi lines revealed that DLA2 is involved in the synthesis of both D1 and acetyl-CoA. Gel filtration analyses demonstrated an RNP complex containing DLA2 and the chloroplast psbA mRNA specifically in cells metabolizing acetate. An intrinsic RNA binding activity of DLA2 was confirmed by in vitro RNA binding assays. Results of fluorescence microscopy and subcellular fractionation experiments support a role of DLA2 in acetate-dependent localization of the psbA mRNA to a translation zone within the chloroplast. Reciprocally, the activity of the cpPDC was specifically affected by binding of psbA mRNA. Beyond that, in silico analysis and in vitro RNA binding studies using recombinant proteins support the possibility that RNA binding is an ancient feature of dihydrolipoamide acetyltransferases. Our results suggest a regulatory function of DLA2 in response to growth on reduced carbon energy sources. This raises the intriguing possibility that this regulation functions to coordinate the synthesis of lipids and proteins for the biogenesis of photosynthetic membranes.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
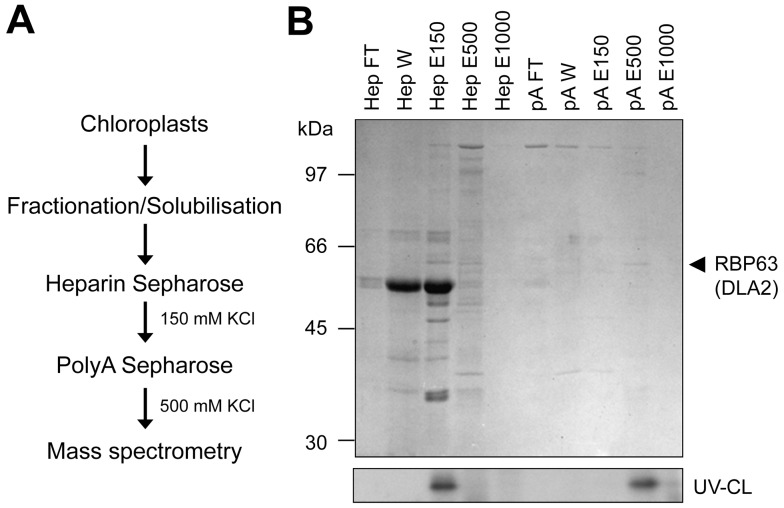

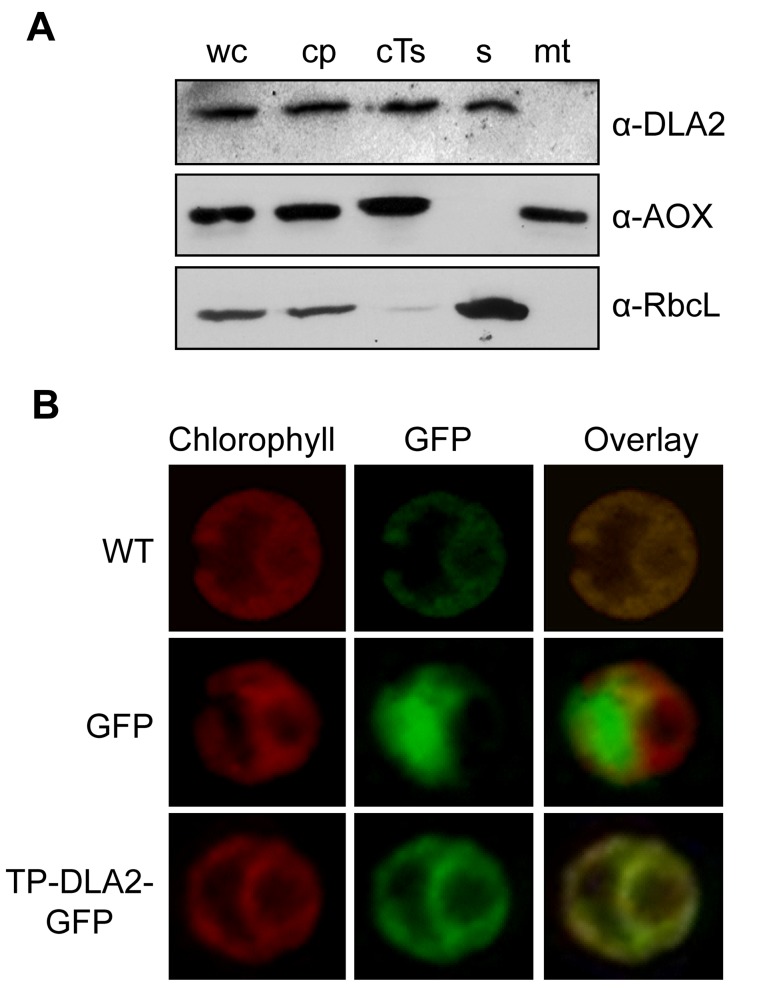
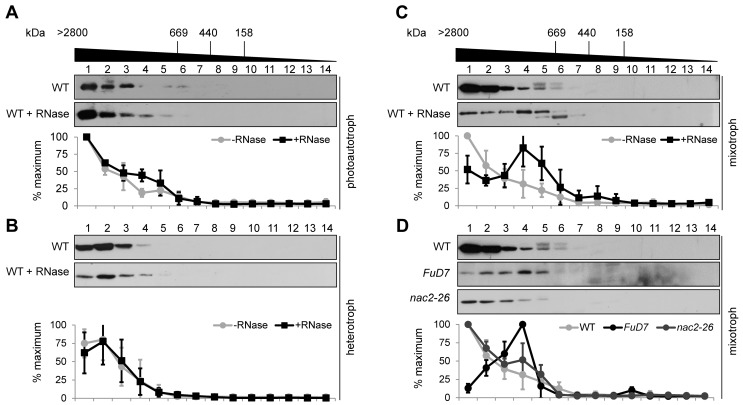
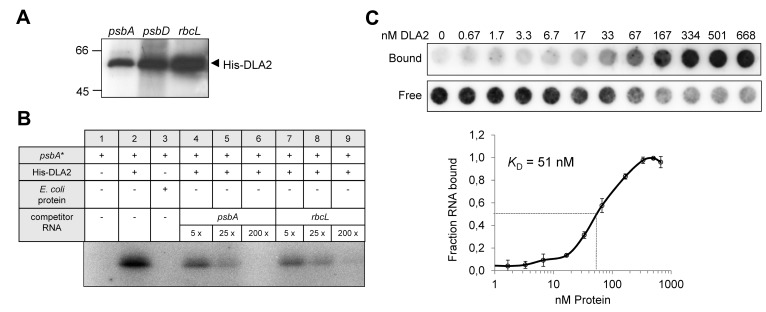
References
-
- McKnight SL (2010) On getting there from here. Science 330: 1338–1339. - PubMed
-
- Wellen KE, Thompson CB (2012) A two-way street: reciprocal regulation of metabolism and signalling. Nat Rev Mol Cell Biol 13: 270–276. - PubMed
-
- Xing S, Poirier Y (2012) The protein acetylome and the regulation of metabolism. Trends Plant Sci 17: 423–430. - PubMed
-
- Bhardwaj A, Wilkinson MF (2005) A metabolic enzyme doing double duty as a transcription factor. Bioessays 27: 467–471. - PubMed
-
- Cieśla J (2006) Metabolic enzymes that bind RNA: yet another level of cellular regulatory network? Acta Biochim Pol 53: 11–32. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
