

mTOR regulates tau phosphorylation and degradation: implications for Alzheimer's disease and other tauopathies
- PMID: 23425014
- PMCID: PMC3655115
- DOI: 10.1111/acel.12057
mTOR regulates tau phosphorylation and degradation: implications for Alzheimer's disease and other tauopathies
Abstract
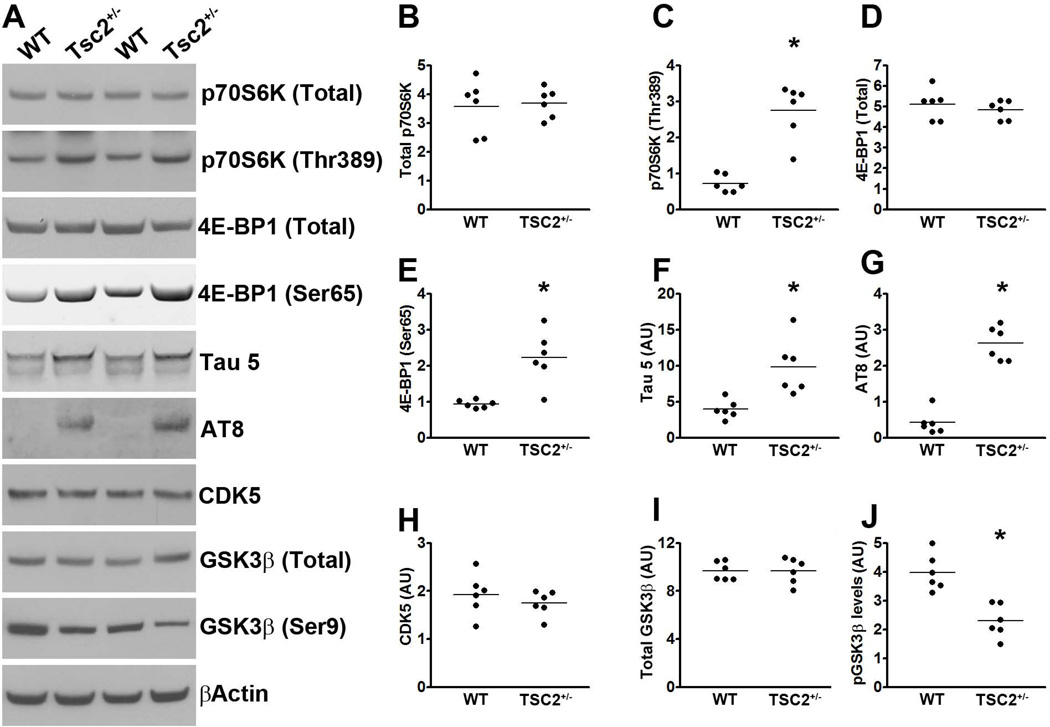
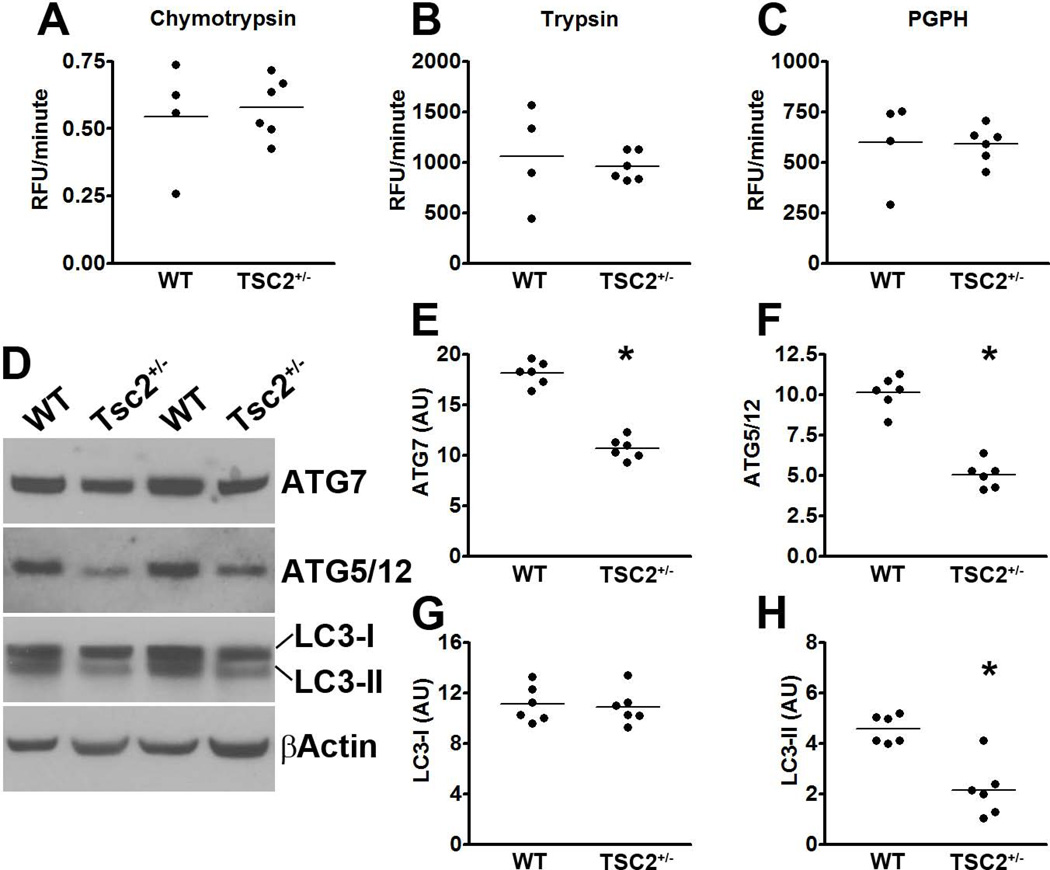
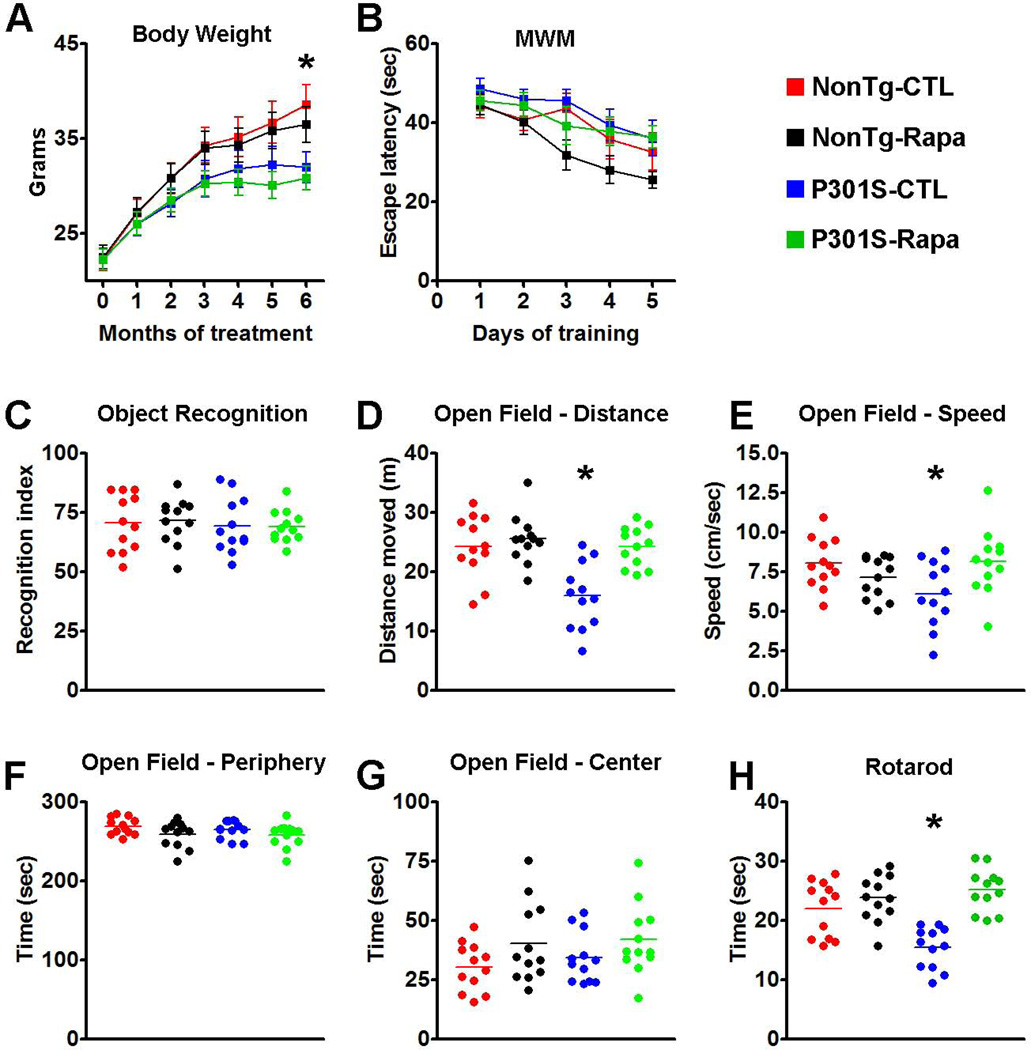
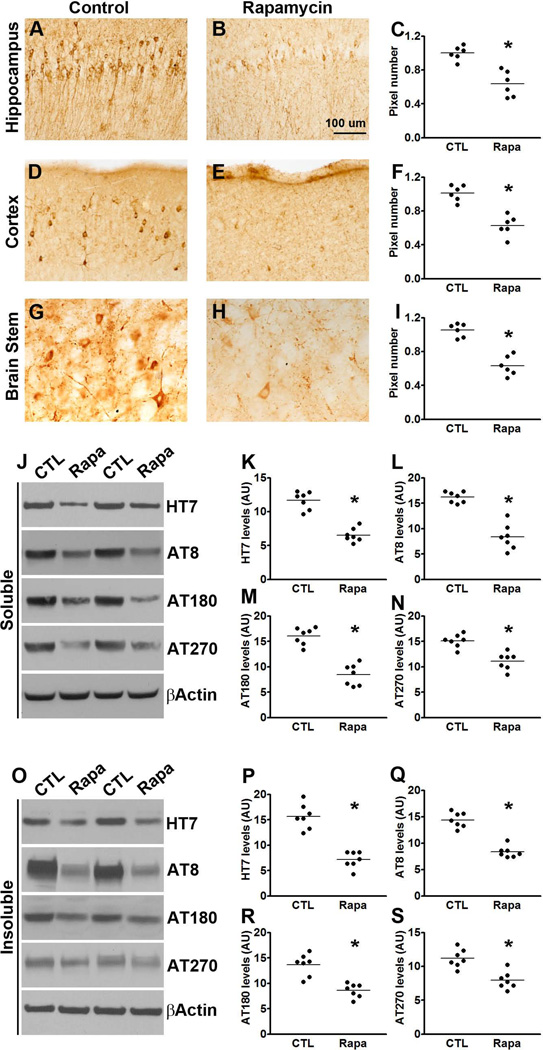
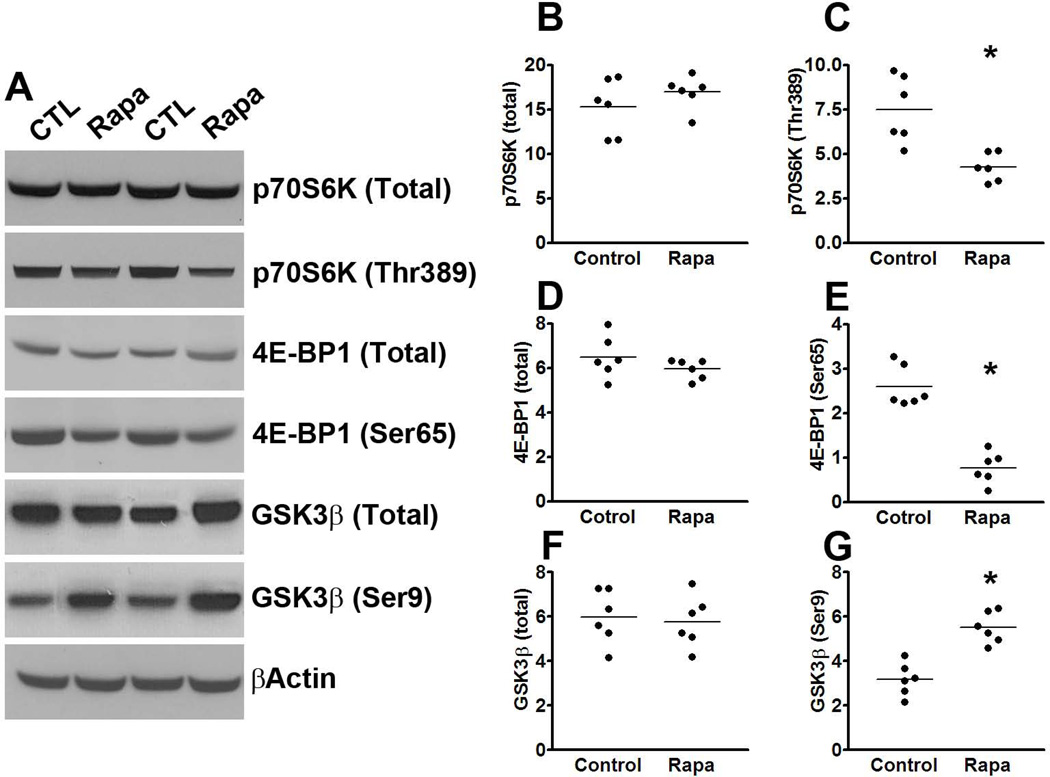
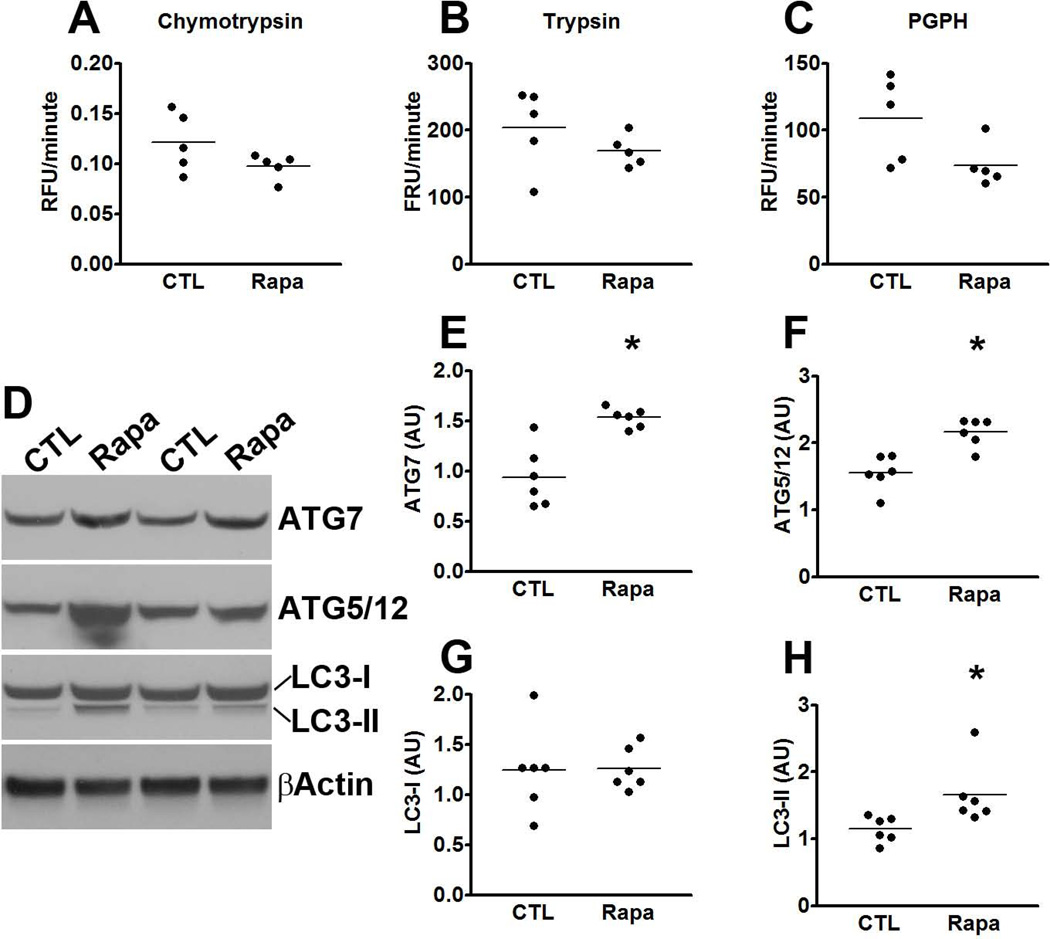
Accumulation of tau is a critical event in several neurodegenerative disorders, collectively known as tauopathies, which include Alzheimer's disease and frontotemporal dementia. Pathological tau is hyperphosphorylated and aggregates to form neurofibrillary tangles. The molecular mechanisms leading to tau accumulation remain unclear and more needs to be done to elucidate them. Age is a major risk factor for all tauopathies, suggesting that molecular changes contributing to the aging process may facilitate tau accumulation and represent common mechanisms across different tauopathies. Here, we use multiple animal models and complementary genetic and pharmacological approaches to show that the mammalian target of rapamycin (mTOR) regulates tau phosphorylation and degradation. Specifically, we show that genetically increasing mTOR activity elevates endogenous mouse tau levels and phosphorylation. Complementary to it, we further demonstrate that pharmacologically reducing mTOR signaling with rapamycin ameliorates tau pathology and the associated behavioral deficits in a mouse model overexpressing mutant human tau. Mechanistically, we provide compelling evidence that the association between mTOR and tau is linked to GSK3β and autophagy function. In summary, we show that increasing mTOR signaling facilitates tau pathology, while reducing mTOR signaling ameliorates tau pathology. Given the overwhelming evidence that reducing mTOR signaling increases lifespan and healthspan, the data presented here have profound clinical implications for aging and tauopathies and provide the molecular basis for how aging may contribute to tau pathology. Additionally, these results provide preclinical data indicating that reducing mTOR signaling may be a valid therapeutic approach for tauopathies.
© 2013 John Wiley & Sons Ltd and the Anatomical Society.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
