

Cigarette smoke mediates epigenetic repression of miR-487b during pulmonary carcinogenesis
- PMID: 23426183
- PMCID: PMC3582115
- DOI: 10.1172/JCI61271
Cigarette smoke mediates epigenetic repression of miR-487b during pulmonary carcinogenesis
Abstract
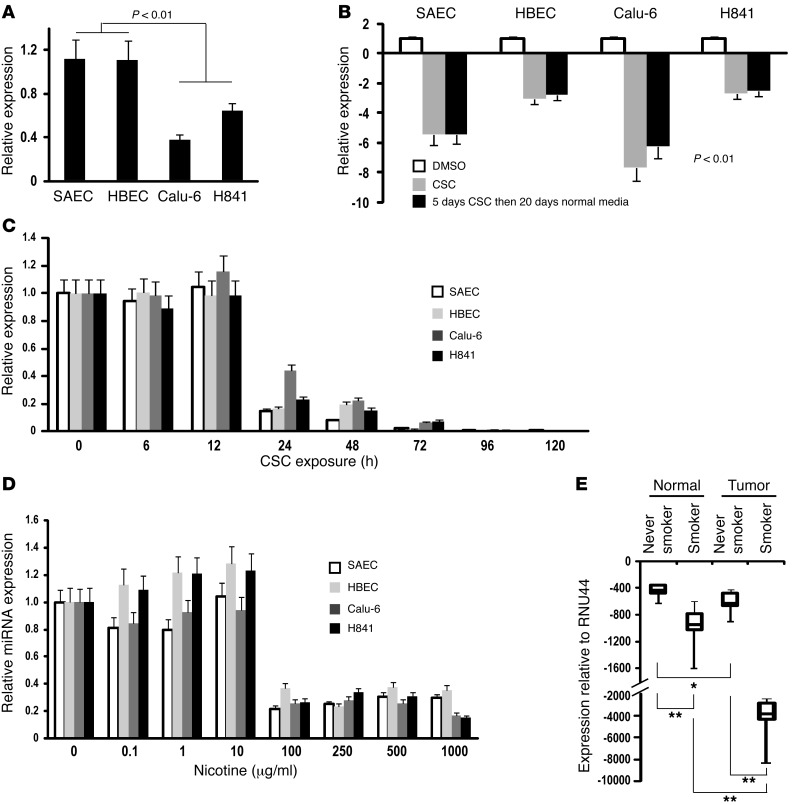
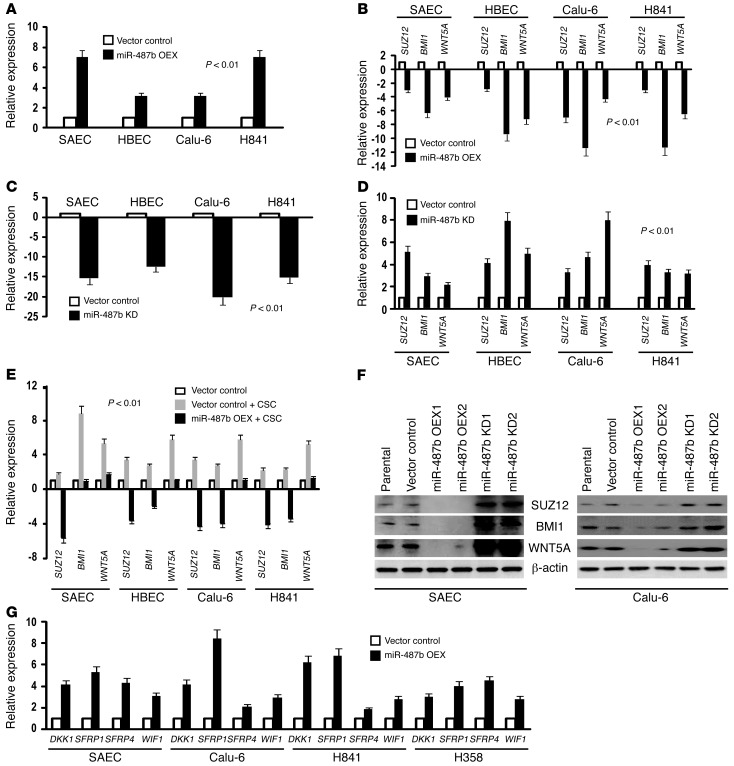
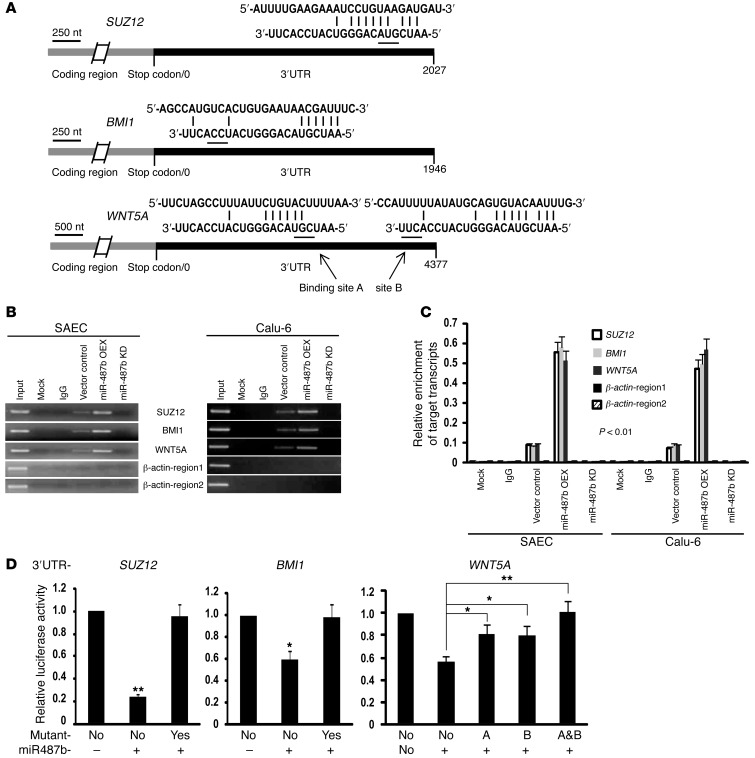
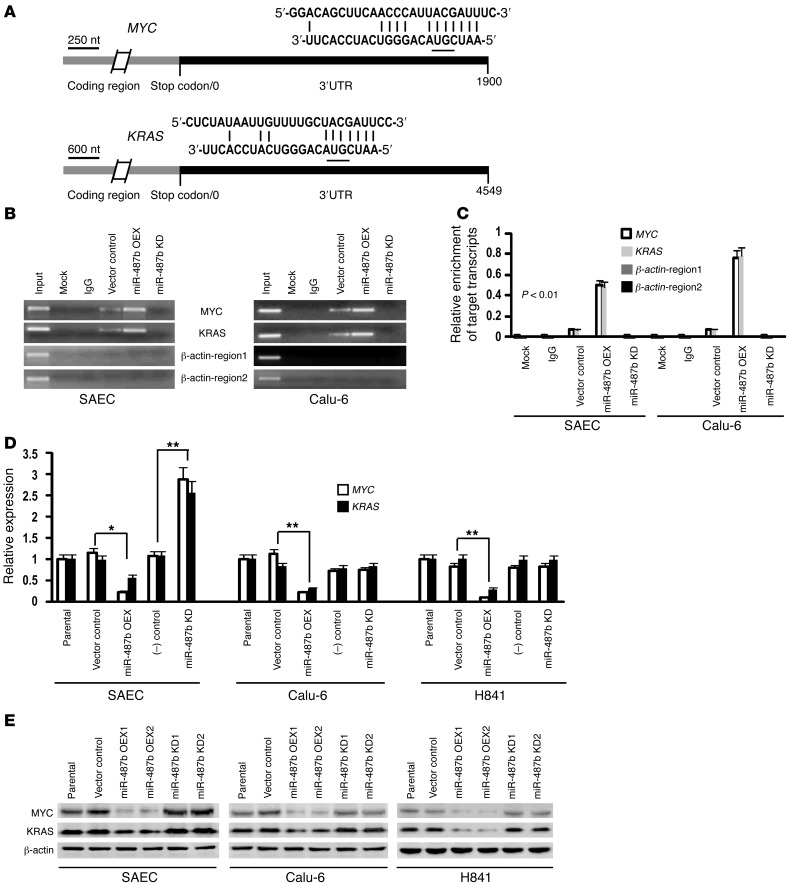
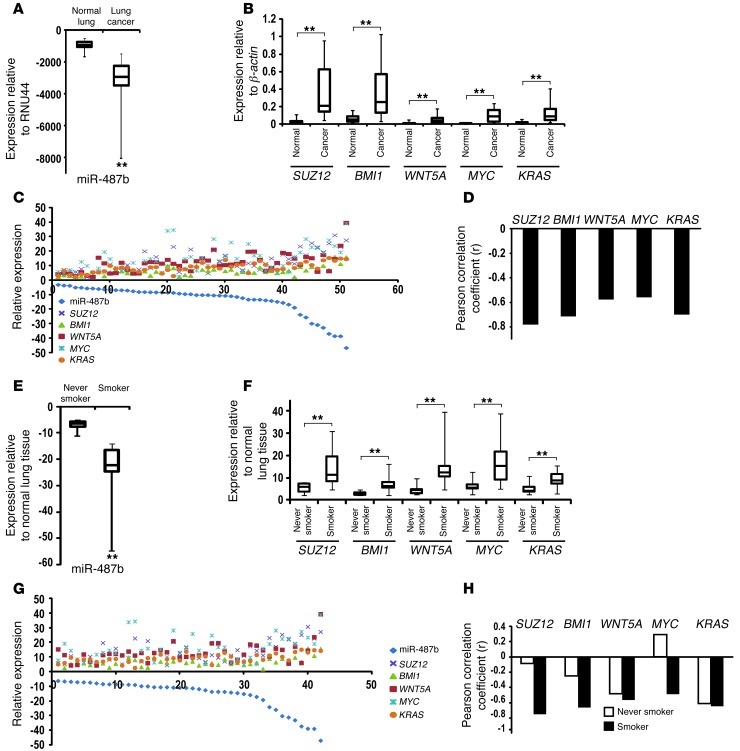
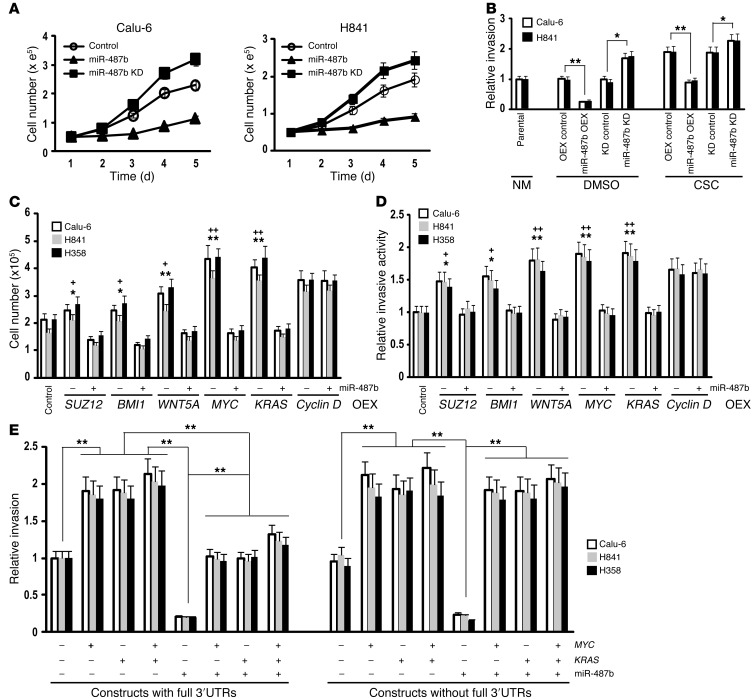
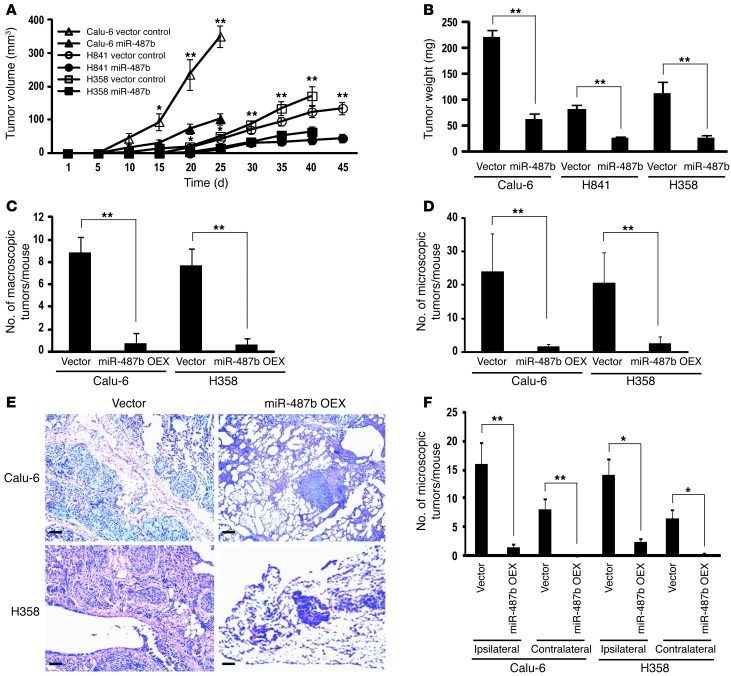
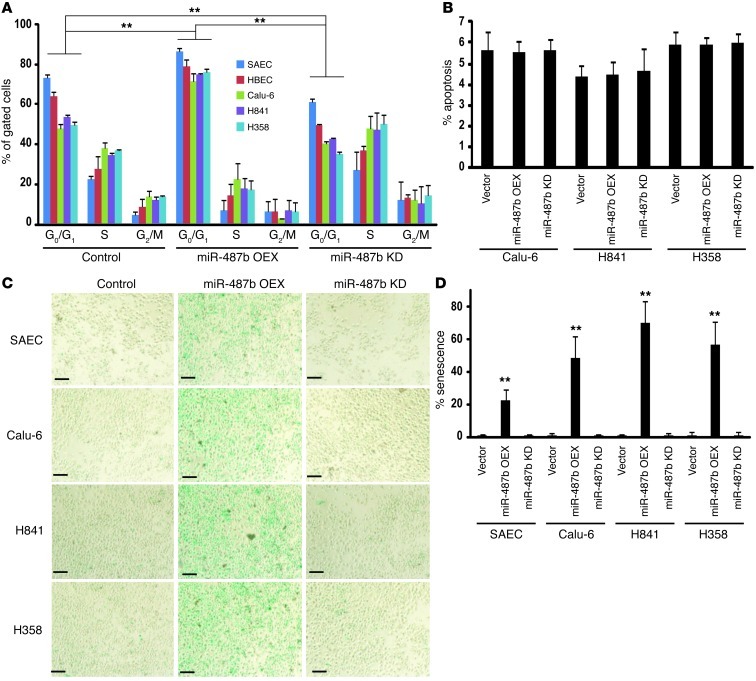
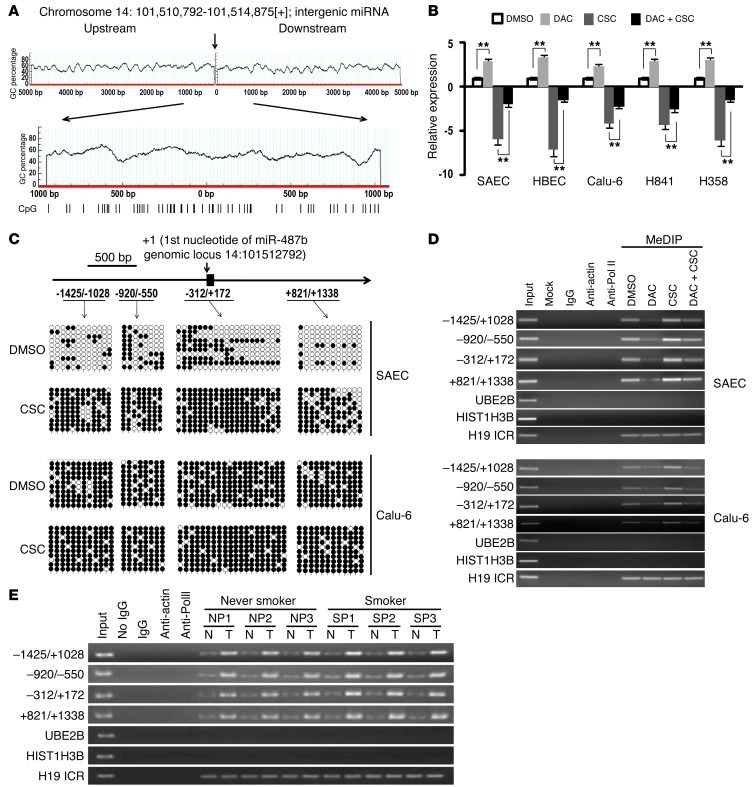
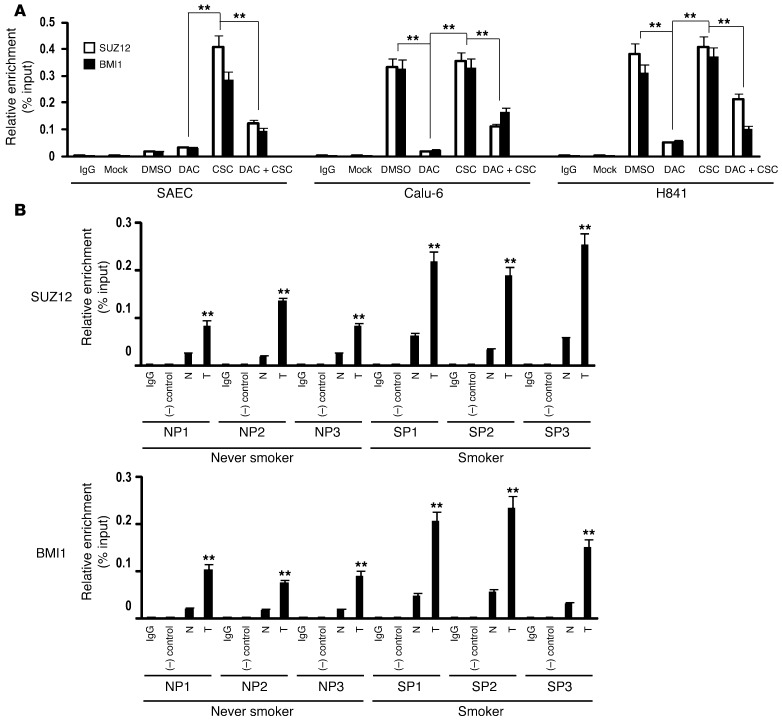
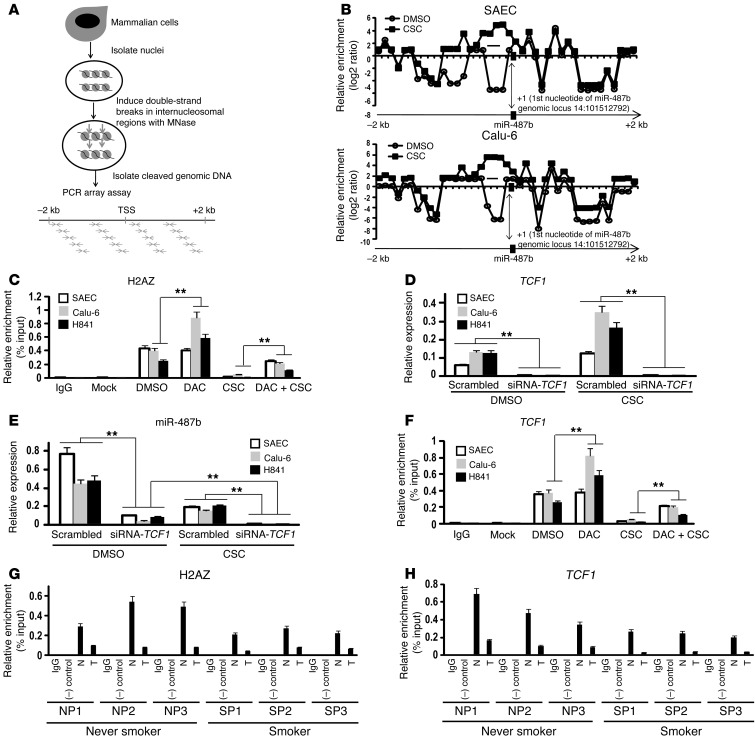
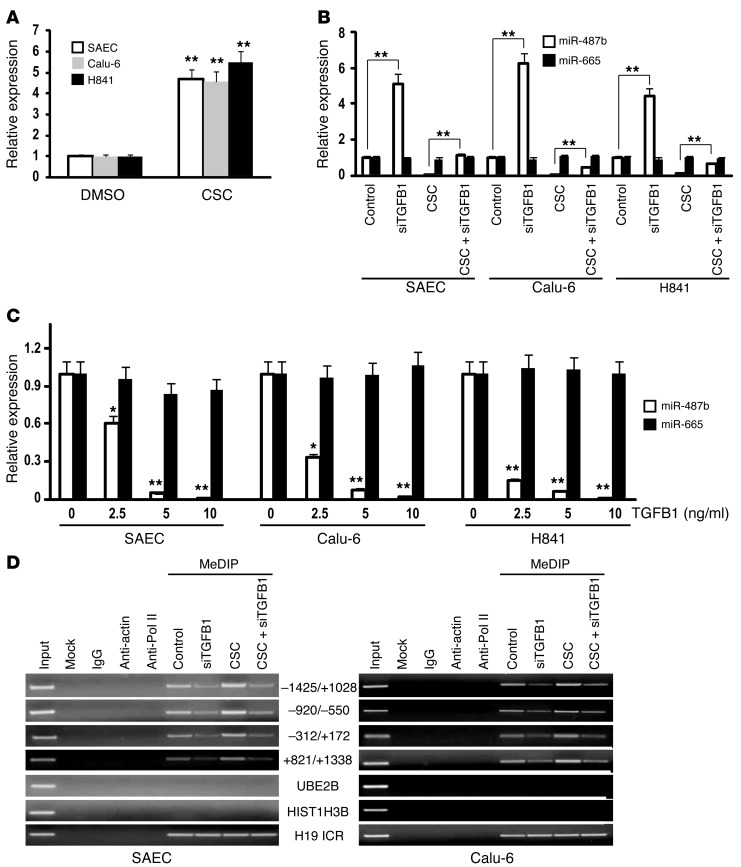
MicroRNAs are critical mediators of stem cell pluripotency, differentiation, and malignancy. Limited information exists regarding microRNA alterations that facilitate initiation and progression of human lung cancers. In this study, array techniques were used to evaluate microRNA expression in normal human respiratory epithelia and lung cancer cells cultured in the presence or absence of cigarette smoke condensate (CSC). Under relevant exposure conditions, CSC significantly repressed miR-487b. Subsequent experiments demonstrated that miR-487b directly targeted SUZ12, BMI1, WNT5A, MYC, and KRAS. Repression of miR-487b correlated with overexpression of these targets in primary lung cancers and coincided with DNA methylation, de novo nucleosome occupancy, and decreased H2AZ and TCF1 levels within the miR-487b genomic locus. Deoxy-azacytidine derepressed miR-487b and attenuated CSC-mediated silencing of miR-487b. Constitutive expression of miR-487b abrogated Wnt signaling, inhibited in vitro proliferation and invasion of lung cancer cells mediated by CSC or overexpression of miR-487b targets, and decreased growth and metastatic potential of lung cancer cells in vivo. Collectively, these findings indicate that miR-487b is a tumor suppressor microRNA silenced by epigenetic mechanisms during tobacco-induced pulmonary carcinogenesis and suggest that DNA demethylating agents may be useful for activating miR-487b for lung cancer therapy.
Figures
References
-
- Malumbres M. miRNAs and cancer: an epigenetics view [published online ahead of print July 4, 2012]. Mol Aspects Med. doi: 10.1016/j.mam.2012.06.005. doi: 10.1016/j.mam.2012.06.005 . - DOI - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
