

Evidence for context-dependent complementarity of non-Shine-Dalgarno ribosome binding sites to Escherichia coli rRNA
- PMID: 23427812
- PMCID: PMC3666857
- DOI: 10.1021/cb3005726
Evidence for context-dependent complementarity of non-Shine-Dalgarno ribosome binding sites to Escherichia coli rRNA
Abstract
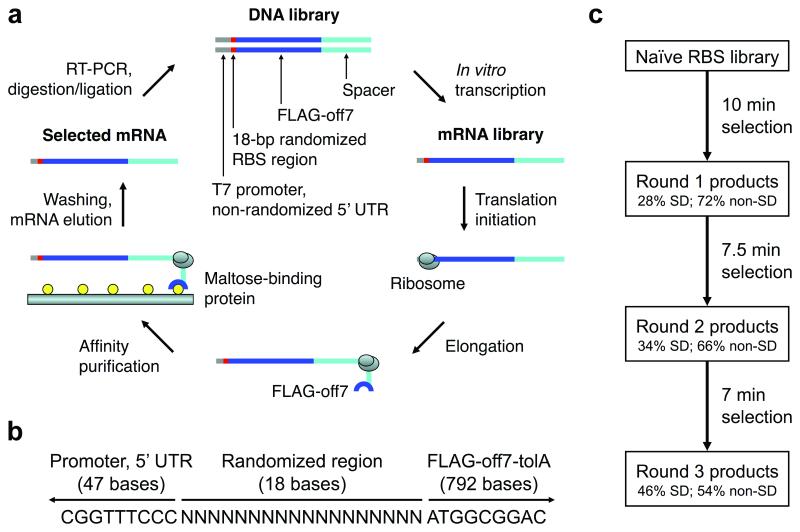
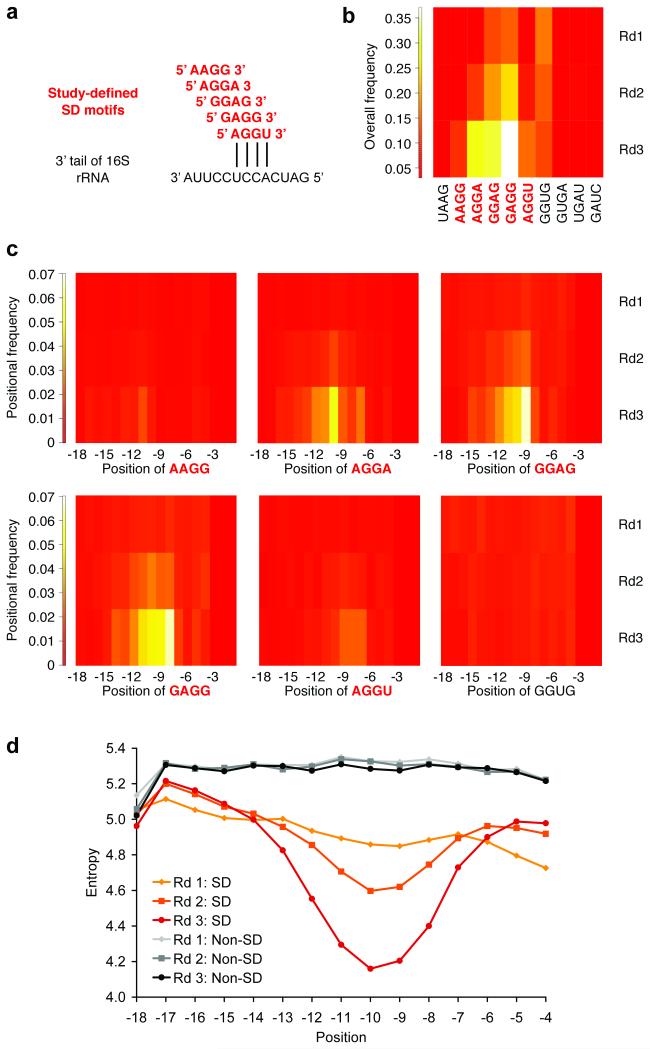
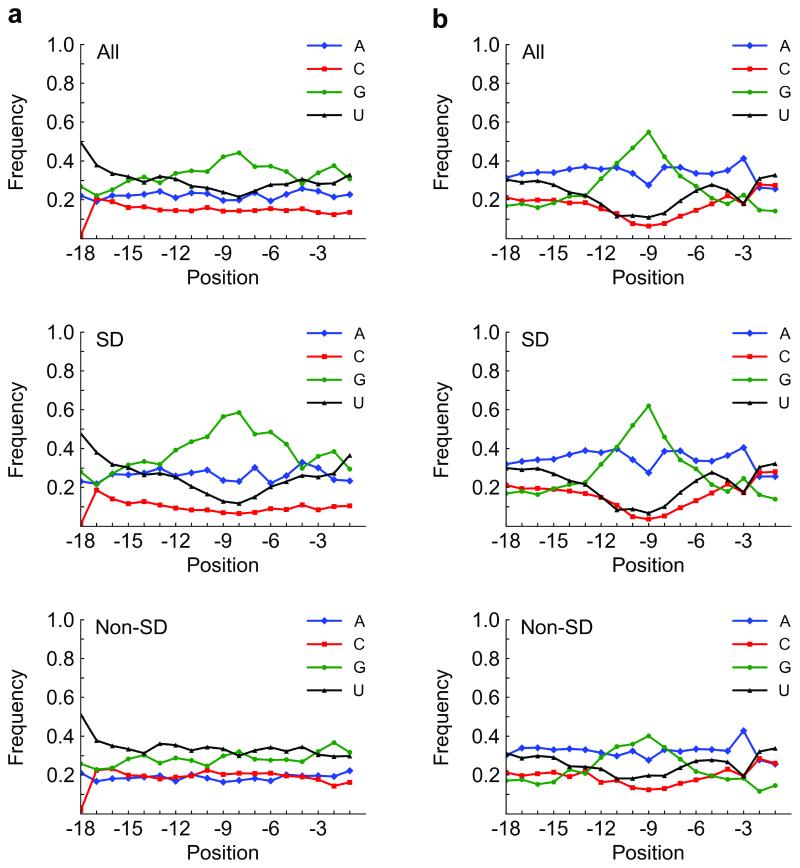
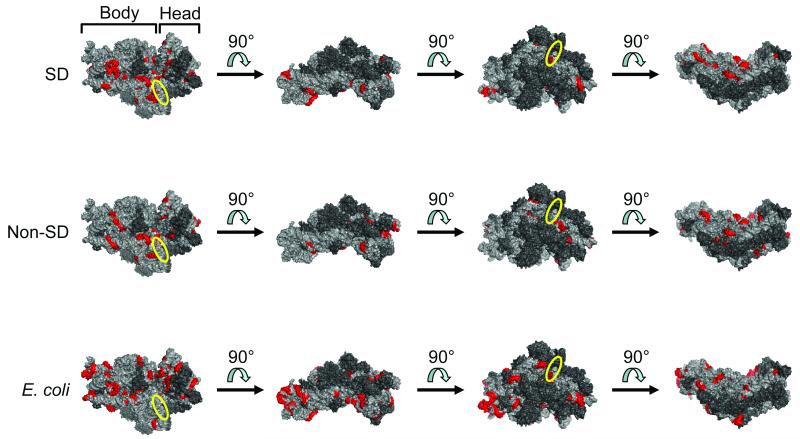
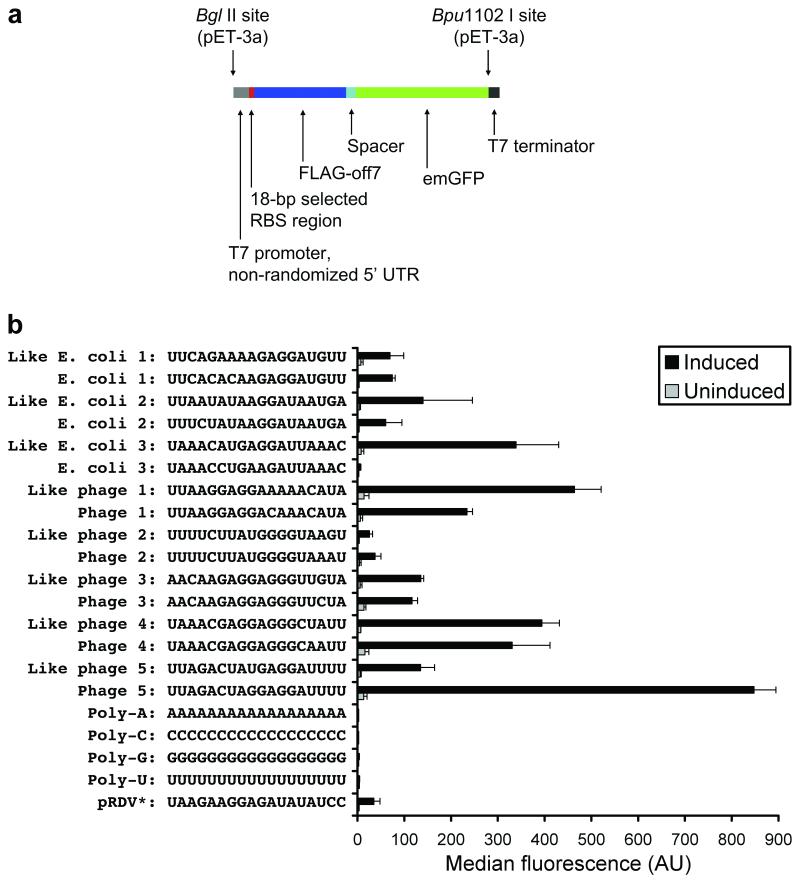
While the ribosome has evolved to function in complex intracellular environments, these contexts do not easily allow for the study of its inherent capabilities. We have used a synthetic, well-defined Escherichia coli (E. coli)-based translation system in conjunction with ribosome display, a powerful in vitro selection method, to identify ribosome binding sites (RBSs) that can promote the efficient translation of messenger RNAs (mRNAs) with a leader length representative of natural E. coli mRNAs. In previous work, we used a longer leader sequence and unexpectedly recovered highly efficient cytosine-rich sequences with complementarity to the 16S ribosomal RNA (rRNA) and similarity to eukaryotic RBSs. In the current study, Shine-Dalgarno (SD) sequences were prevalent, but non-SD sequences were also heavily enriched and were dominated by novel guanine- and uracil-rich motifs that showed statistically significant complementarity to the 16S rRNA. Additionally, only SD motifs exhibited position-dependent decreases in sequence entropy, indicating that non-SD motifs likely operate by increasing the local concentration of ribosomes in the vicinity of the start codon, rather than by a position-dependent mechanism. These results further support the putative generality of mRNA-rRNA complementarity in facilitating mRNA translation but also suggest that context (e.g., leader length and composition) dictates the specific subset of possible RBSs that are used for efficient translation of a given transcript.
Figures
References
-
- Chang B, Halgamuge S, Tang S-L. Analysis of SD sequences in completed microbial genomes: non-SD-led genes are as common as SD-led genes. Gene. 2006;373:90–99. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
