

Tristetraprolin (TTP): interactions with mRNA and proteins, and current thoughts on mechanisms of action
- PMID: 23428348
- PMCID: PMC3752887
- DOI: 10.1016/j.bbagrm.2013.02.003
Tristetraprolin (TTP): interactions with mRNA and proteins, and current thoughts on mechanisms of action
Abstract
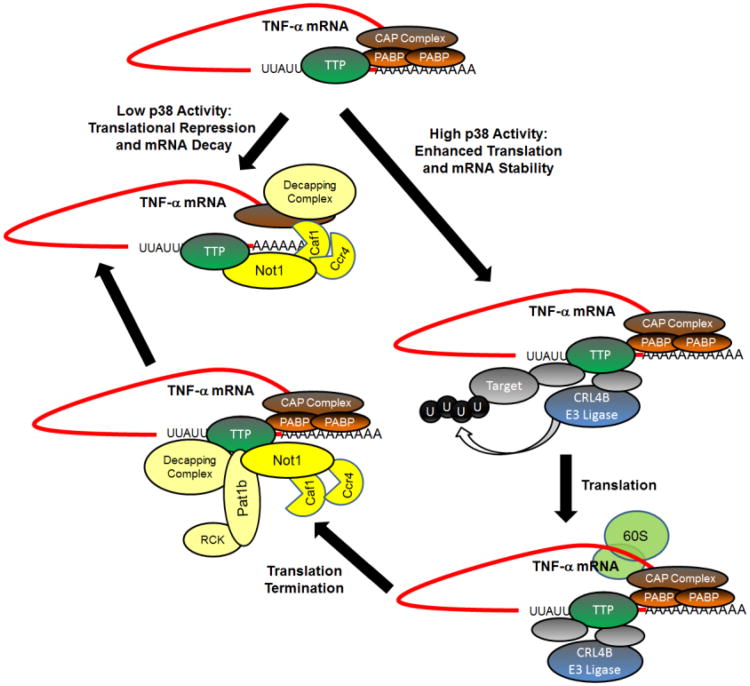
Changes in mRNA stability and translation are critical control points in the regulation of gene expression, particularly genes encoding growth factors, inflammatory mediators, and proto-oncogenes. Adenosine and uridine (AU)-rich elements (ARE), often located in the 3' untranslated regions (3'UTR) of mRNAs, are known to target transcripts for rapid decay. They are also involved in the regulation of mRNA stability and translation in response to extracellular cues. This review focuses on one of the best characterized ARE binding proteins, tristetraprolin (TTP), the founding member of a small family of CCCH tandem zinc finger proteins. In this survey, we have reviewed the current status of TTP interactions with mRNA and proteins, and discussed current thinking about TTP's mechanism of action to promote mRNA decay. We also review the proposed regulation of TTP's functions by phosphorylation. Finally, we have discussed emerging evidence for TTP operating as a translational regulator. This article is part of a Special Issue entitled: RNA Decay mechanisms.
Published by Elsevier B.V.
Figures

References
-
- Benjamin D, Moroni C. mRNA stability and cancer: an emerging link? Expert Opin Biol Ther. 2007;7:1515–1529. - PubMed
-
- Hitti E, Khabar KS. Sequence variations affecting AU-rich element function and disease. Front Biosci. 2012;17:1846–1860. - PubMed
-
- Schott J, Stoecklin G. Networks controlling mRNA decay in the immune system. Wiley Interdiscip Rev RNA. 2010;1:432–456. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
