

Photoperiodic regulation of the sucrose transporter StSUT4 affects the expression of circadian-regulated genes and ethylene production
- PMID: 23429841
- PMCID: PMC3576705
- DOI: 10.3389/fpls.2013.00026
Photoperiodic regulation of the sucrose transporter StSUT4 affects the expression of circadian-regulated genes and ethylene production
Abstract
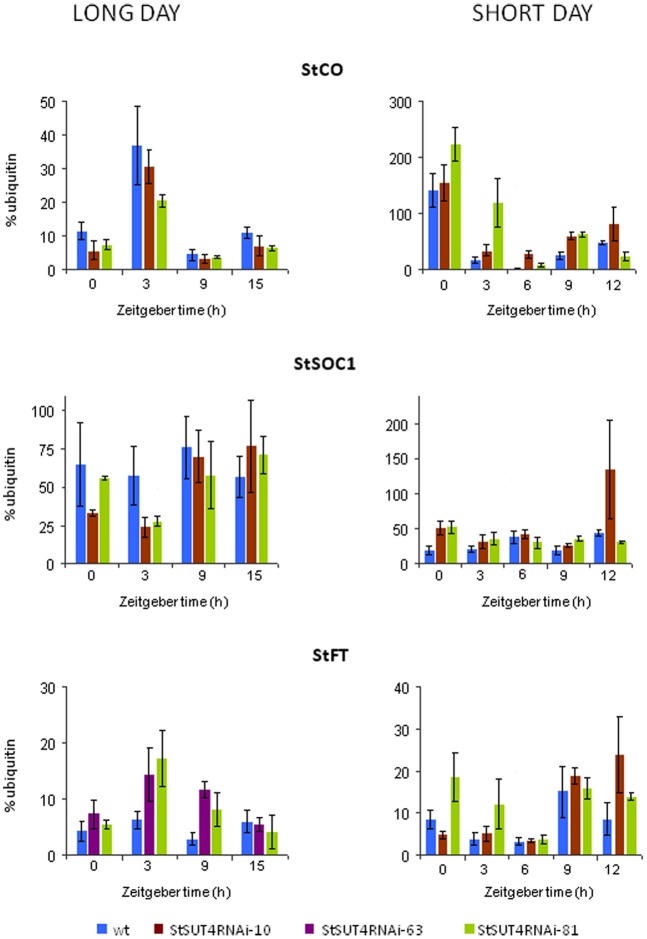
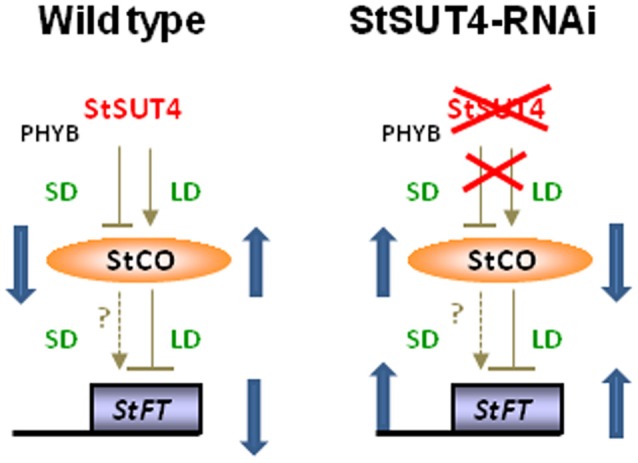
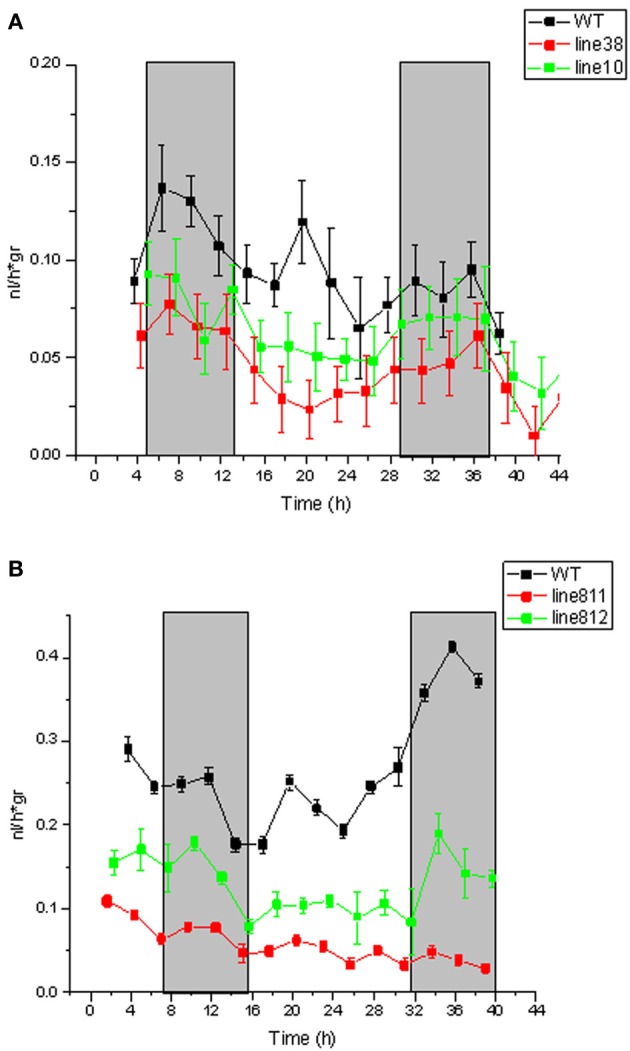
Several recent publications reported different subcellular localization of the sucrose transporters belonging to the SUT4 subfamily. The physiological function of the SUT4 sucrose transporters requires clarification, because down-regulation of the members of the SUT4 clade had different effects in rice, poplar, and potato. Here, we provide new data for the localization and function of the Solanaceous StSUT4 protein, further elucidating involvement in the onset of flowering, tuberization and in the shade avoidance syndrome of potato plants. Induction of an early flowering and a tuberization in the SUT4-inhibited potato plants correlates with increased sucrose export from leaves and increased sucrose and starch accumulation in terminal sink organs, such as developing tubers. SUT4 affects expression of the enzymes involved in gibberellin and ethylene biosynthesis, as well as the rate of ethylene biosynthesis in potato. In the SUT4-inhibited plants, the ethylene production no longer follows a diurnal rhythm. Thus it was concluded that StSUT4 controls circadian gene expression, potentially by regulating sucrose export from leaves. Furthermore, SUT4 expression affects clock-regulated genes such as StFT, StSOC1, and StCO, which might be also involved in a photoperiod-dependent tuberization. A model is proposed in which StSUT4 controls a phloem-mobile signaling molecule generated in leaves, which together with enhanced sucrose export affects developmental switches in apical meristems. SUT4 seems to link photoreceptor-perceived information about the light quality and day length with phytohormone biosynthesis and the expression of circadian-regulated genes.
Keywords: ethylene; flowering; shade avoidance syndrome; sucrose transport.
Figures
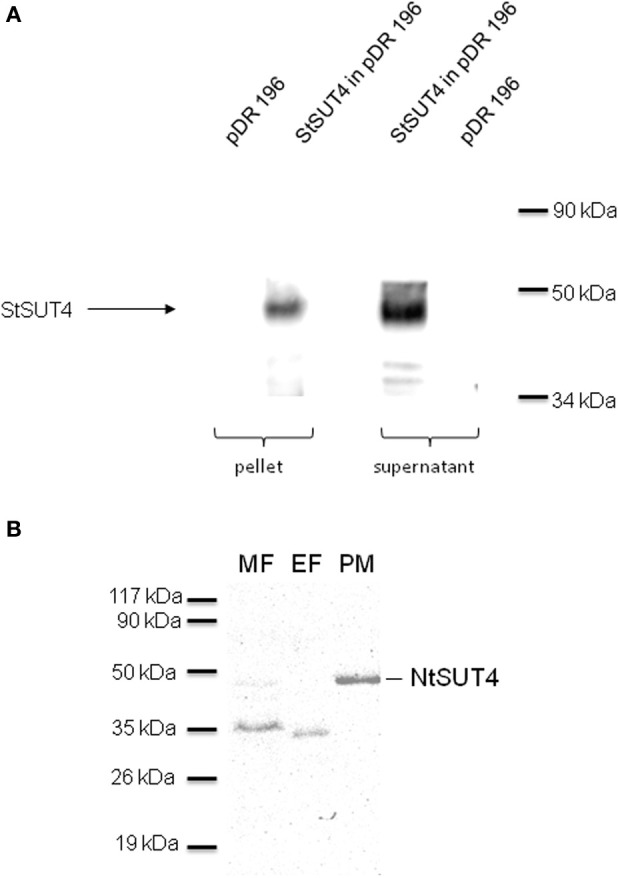
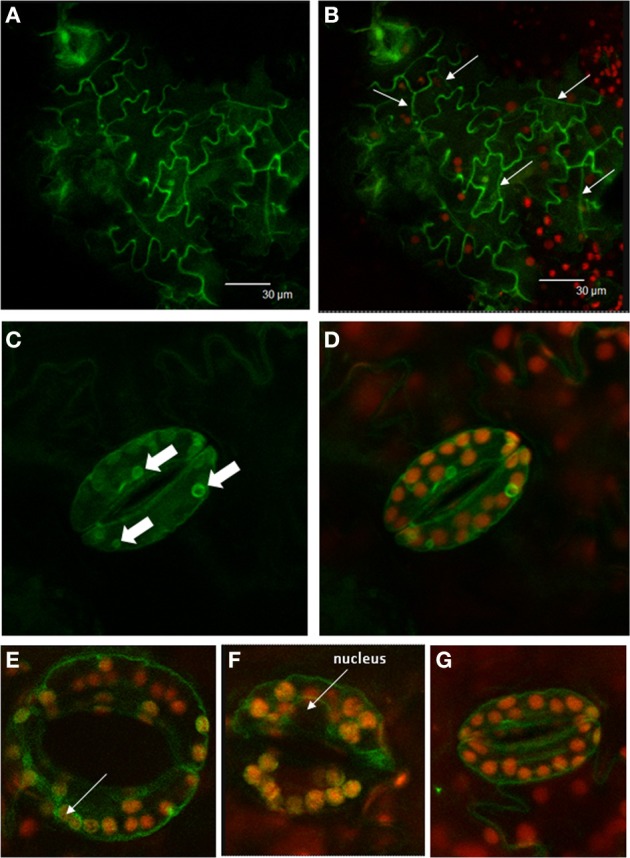
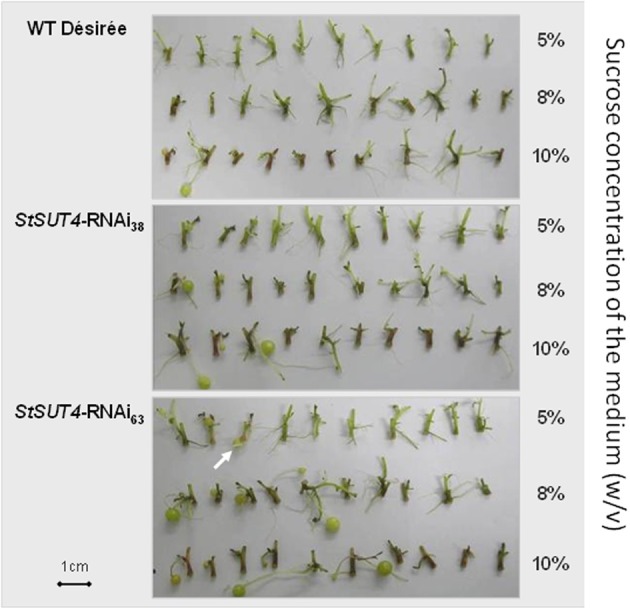
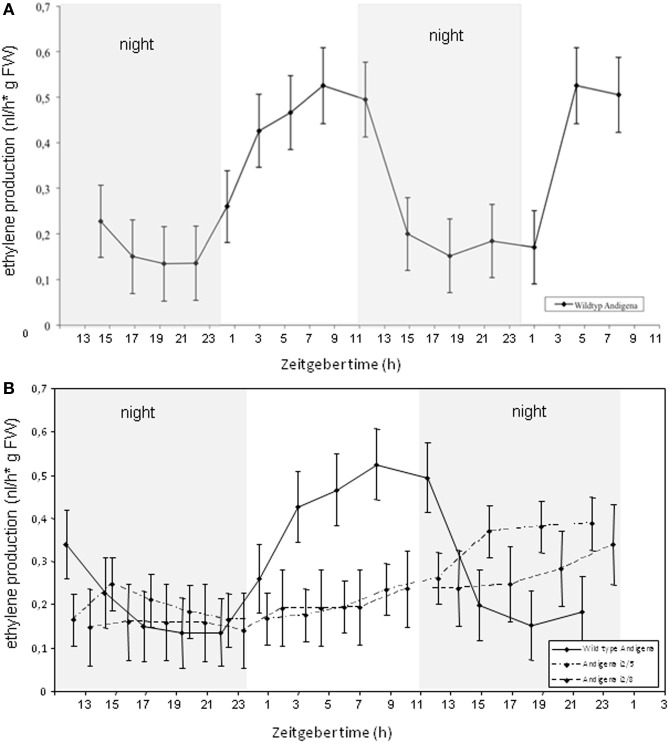
References
-
- Achard P., Baghour M., Chapple A., Hedden P., Van Der Straeten D., Genschik P., et al. (2007). The plant stress hormone ethylene controls floral transition via DELLA-dependent regulation of floral meristem-identity genes. Proc. Natl. Acad. Sci. U.S.A. 104, 6484–6489 10.1073/pnas.0610717104 - DOI - PMC - PubMed
-
- Alders A. W. C. (1987). Marine Refrigeration Manual. Rotterdam: Rotterdam Marine Chartering Agents
LinkOut - more resources
Full Text Sources
Other Literature Sources