

Variable electron transfer pathways in an amphibian cryptochrome: tryptophan versus tyrosine-based radical pairs
- PMID: 23430261
- PMCID: PMC3610996
- DOI: 10.1074/jbc.M112.417725
Variable electron transfer pathways in an amphibian cryptochrome: tryptophan versus tyrosine-based radical pairs
Abstract
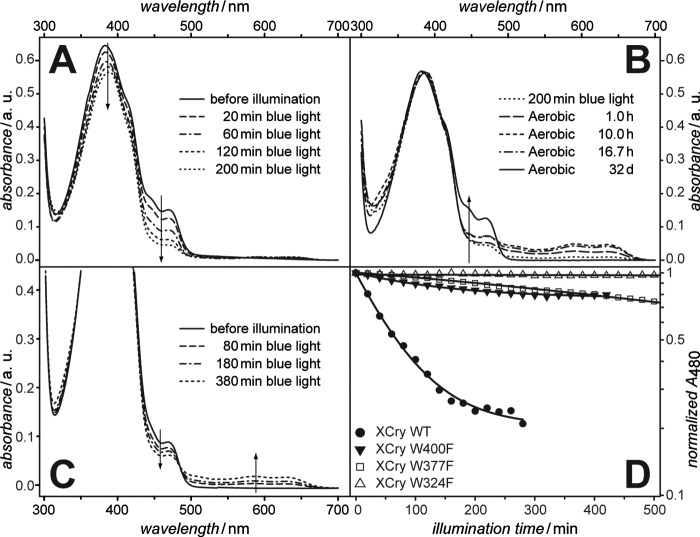
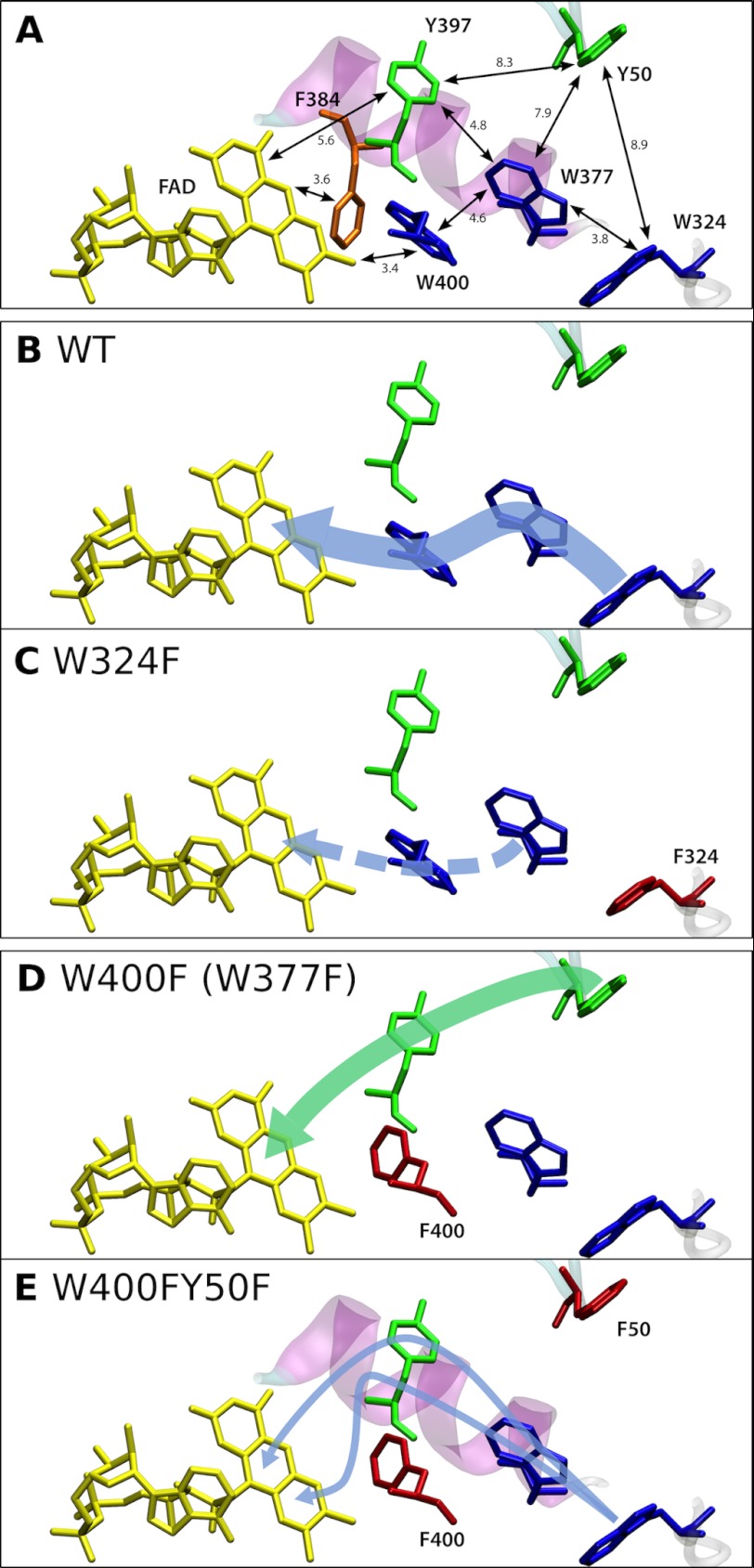
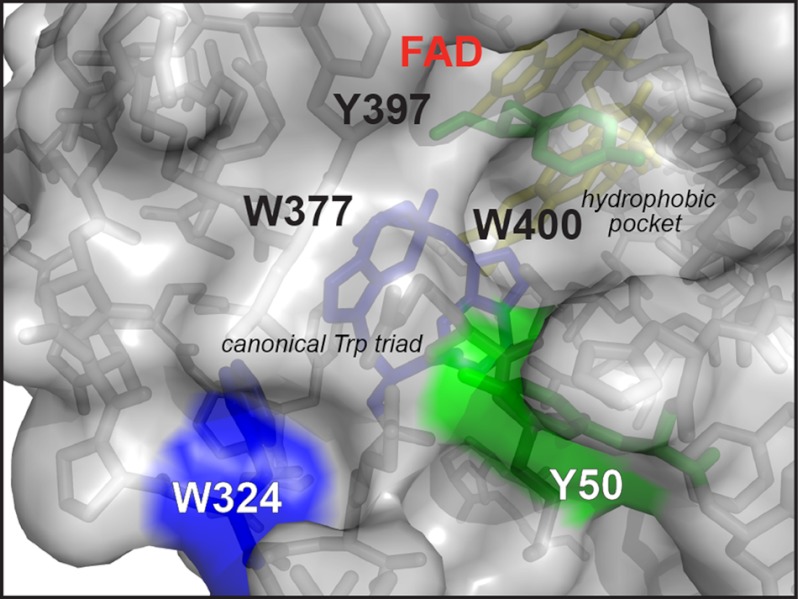
Electron transfer reactions play vital roles in many biological processes. Very often the transfer of charge(s) proceeds stepwise over large distances involving several amino acid residues. By using time-resolved electron paramagnetic resonance and optical spectroscopy, we have studied the mechanism of light-induced reduction of the FAD cofactor of cryptochrome/photolyase family proteins. In this study, we demonstrate that electron abstraction from a nearby amino acid by the excited FAD triggers further electron transfer steps even if the conserved chain of three tryptophans, known to be an effective electron transfer pathway in these proteins, is blocked. Furthermore, we were able to characterize this secondary electron transfer pathway and identify the amino acid partner of the resulting flavin-amino acid radical pair as a tyrosine located at the protein surface. This alternative electron transfer pathway could explain why interrupting the conserved tryptophan triad does not necessarily alter photoreactions of cryptochromes in vivo. Taken together, our results demonstrate that light-induced electron transfer is a robust property of cryptochromes and more intricate than commonly anticipated.
Figures


References
-
- Ahmad M., Cashmore A. R. (1993) HY4 gene of A. thaliana encodes a protein with characteristics of a blue-light photoreceptor. Nature 366, 162–166 - PubMed
-
- Chaves I., Pokorny R., Byrdin M., Hoang N., Ritz T., Brettel K., Essen L. O., van der Horst G. T., Batschauer A., Ahmad M. (2011) The cryptochromes. Blue light photoreceptors in plants and animals. Annu. Rev. Plant Biol. 62, 335–364 - PubMed
-
- Losi A., Gärtner W. (2011) Old chromophores, new photoactivation paradigms, trendy applications. Flavins in blue light-sensing photoreceptors. Photochem. Photobiol. 87, 491–510 - PubMed
-
- Essen L. O. (2006) Photolyases and cryptochromes. Common mechanisms of DNA repair and light-driven signaling? Curr. Opin. Struct. Biol. 16, 51–59 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
