

MITEs in the promoters of effector genes allow prediction of novel virulence genes in Fusarium oxysporum
- PMID: 23432788
- PMCID: PMC3599309
- DOI: 10.1186/1471-2164-14-119
MITEs in the promoters of effector genes allow prediction of novel virulence genes in Fusarium oxysporum
Abstract
Background: The plant-pathogenic fungus Fusarium oxysporum f.sp.lycopersici (Fol) has accessory, lineage-specific (LS) chromosomes that can be transferred horizontally between strains. A single LS chromosome in the Fol4287 reference strain harbors all known Fol effector genes. Transfer of this pathogenicity chromosome confers virulence to a previously non-pathogenic recipient strain. We hypothesize that expression and evolution of effector genes is influenced by their genomic context.
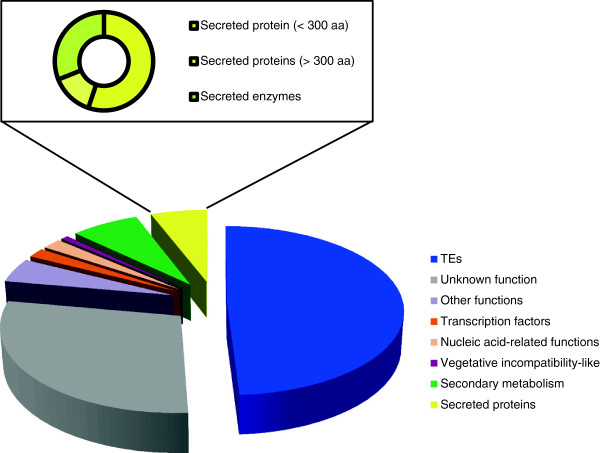
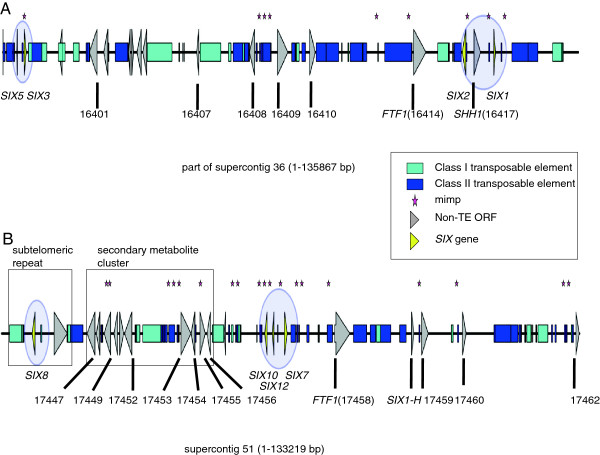
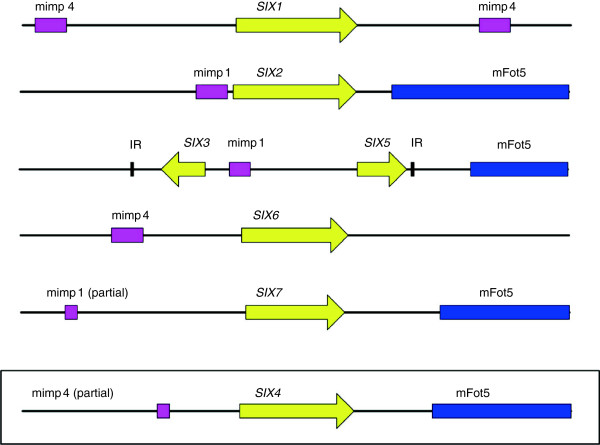
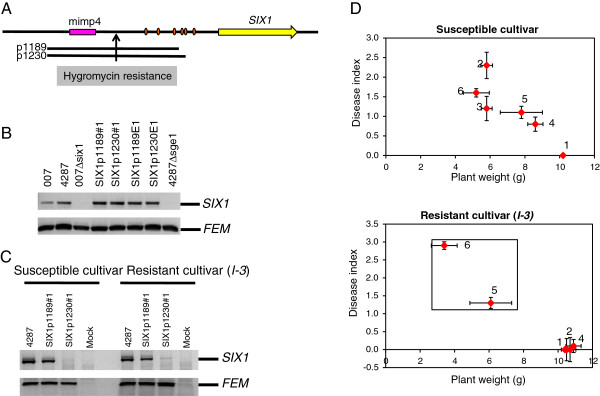
Results: To gain a better understanding of the genomic context of the effector genes, we manually curated the annotated genes on the pathogenicity chromosome and identified and classified transposable elements. Both retro- and DNA transposons are present with no particular overrepresented class. Retrotransposons appear evenly distributed over the chromosome, while DNA transposons tend to concentrate in large chromosomal subregions. In general, genes on the pathogenicity chromosome are dispersed within the repeat landscape. Effector genes are present within subregions enriched for DNA transposons. A miniature Impala (mimp) is always present in their promoters. Although promoter deletion studies of two effector gene loci did not reveal a direct function of the mimp for gene expression, we were able to use proximity to a mimp as a criterion to identify new effector gene candidates. Through xylem sap proteomics we confirmed that several of these candidates encode proteins secreted during plant infection.
Conclusions: Effector genes in Fol reside in characteristic subregions on a pathogenicity chromosome. Their genomic context allowed us to develop a method for the successful identification of novel effector genes. Since our approach is not based on effector gene similarity, but on unique genomic features, it can easily be extended to identify effector genes in Fo strains with different host specificities.
Figures


References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
