

Circadian disruption leads to insulin resistance and obesity
- PMID: 23434278
- PMCID: PMC3595381
- DOI: 10.1016/j.cub.2013.01.048
Circadian disruption leads to insulin resistance and obesity
Abstract
Background: Disruption of circadian (daily) timekeeping enhances the risk of metabolic syndrome, obesity, and type 2 diabetes. While clinical observations have suggested that insulin action is not constant throughout the 24 hr cycle, its magnitude and periodicity have not been assessed. Moreover, when circadian rhythmicity is absent or severely disrupted, it is not known whether insulin action will lock to the peak, nadir, or mean of the normal periodicity of insulin action.
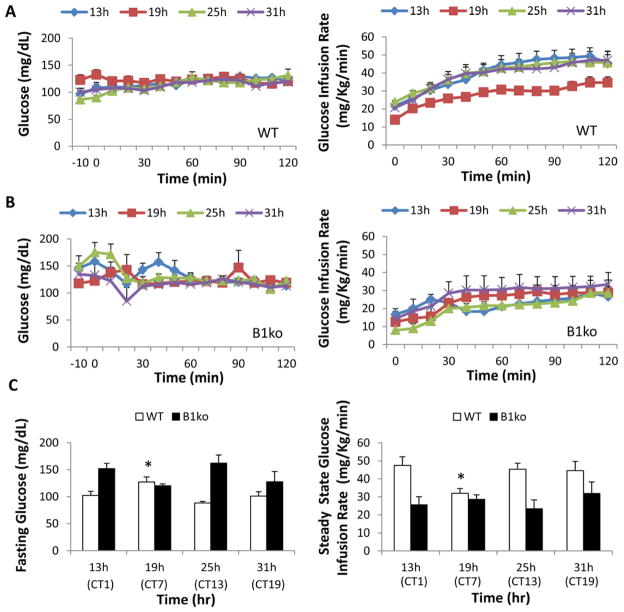
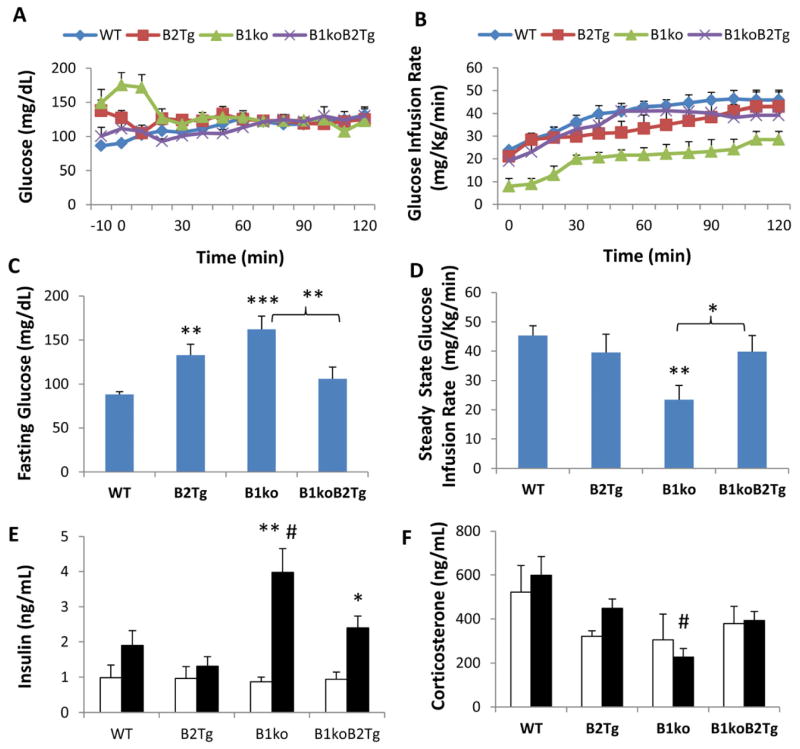
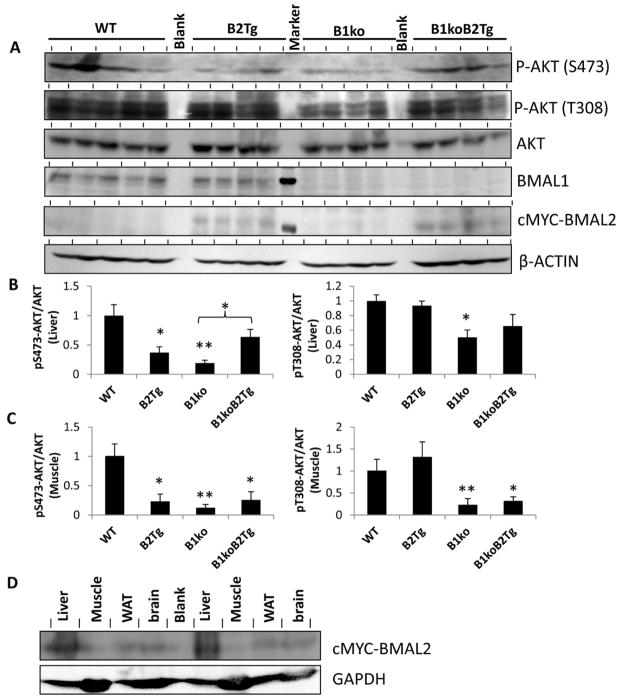
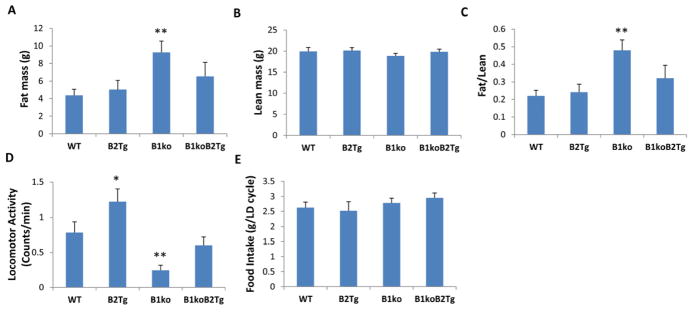
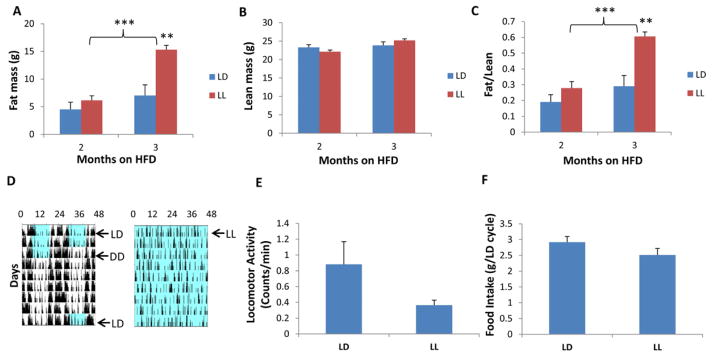
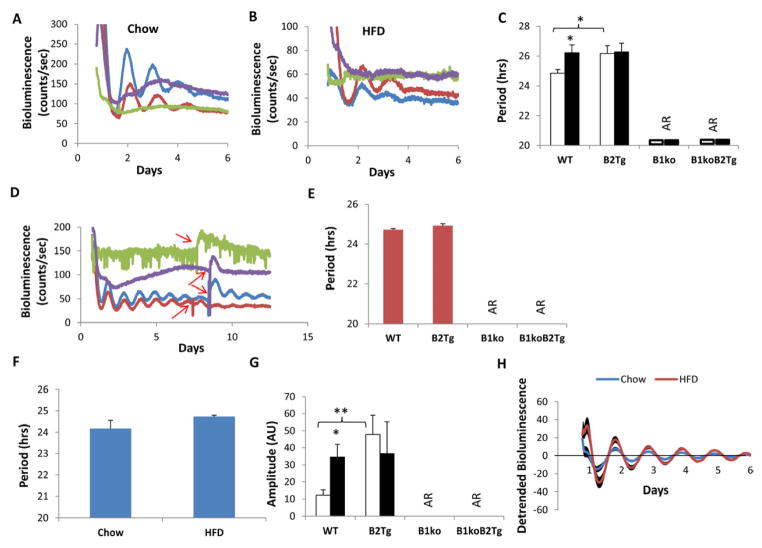
Results: We used hyperinsulinemic-euglycemic clamps to show a bona fide circadian rhythm of insulin action; mice are most resistant to insulin during their daily phase of relative inactivity. Moreover, clock-disrupted Bmal1-knockout mice are locked into the trough of insulin action and lack rhythmicity in insulin action and activity patterns. When rhythmicity is rescued in the Bmal1-knockout mice by expression of the paralogous gene Bmal2, insulin action and activity patterns are restored. When challenged with a high-fat diet, arhythmic mice (either Bmal1-knockout mice or wild-type mice made arhythmic by exposure to constant light) were obese prone. Adipose tissue explants obtained from high-fat-fed mice have their own periodicity that was longer than animals on a chow diet.
Conclusions: This study provides rigorous documentation for a circadian rhythm of insulin action and demonstrates that disturbing the natural rhythmicity of insulin action will disrupt the rhythmic internal environment of insulin sensitive tissue, thereby predisposing the animals to insulin resistance and obesity.
Copyright © 2013 Elsevier Ltd. All rights reserved.
Conflict of interest statement
The authors declare that they have no conflicts of interest.
Figures
References
-
- Dunlap JC, Loros JJ, DeCoursey PJ. Chronobiology: Biological Timekeeping. Sunderland, MA: Sinauer Associates, Inc; 2004.
-
- Gimble JM, Sutton GM, Bunnell BA, Ptitsyn AA, Floyd ZE. Prospective influences of circadian clocks in adipose tissue and metabolism. Nat Rev Endocrinol. 2011;7:98–107. - PubMed
-
- Antunes LC, Levandovski R, Dantas G, Caumo W, Hidalgo MP. Obesity and shift work: chronobiological aspects. Nutr Res Rev. 2010;23:155–68. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
