

The voltage-sensing domain of a phosphatase gates the pore of a potassium channel
- PMID: 23440279
- PMCID: PMC3581695
- DOI: 10.1085/jgp.201210940
The voltage-sensing domain of a phosphatase gates the pore of a potassium channel
Abstract
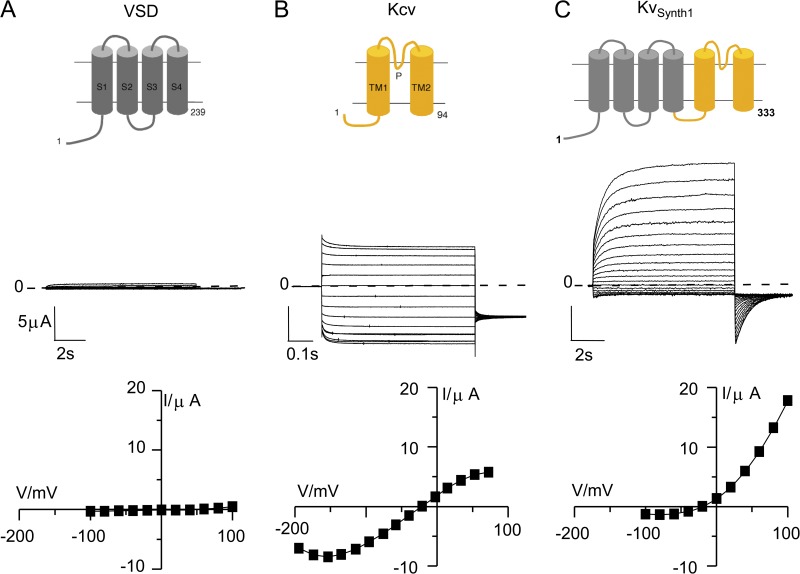
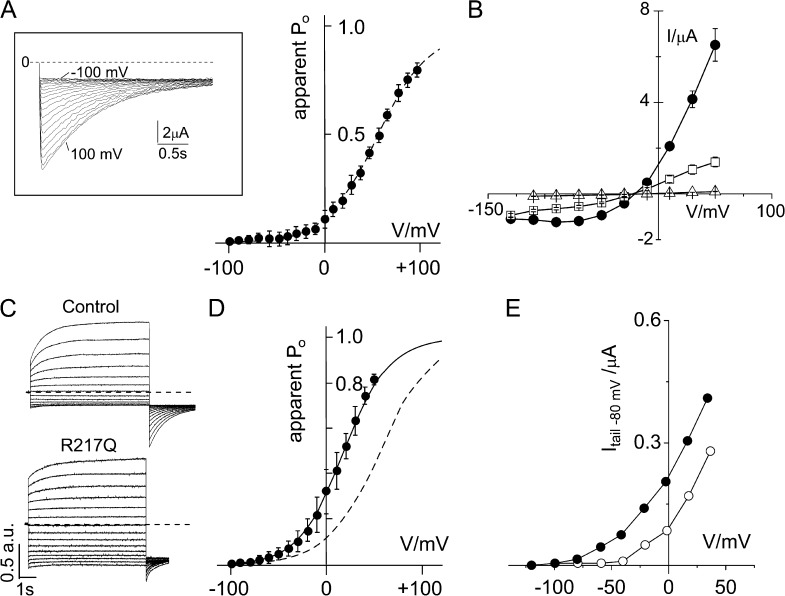
The modular architecture of voltage-gated K(+) (Kv) channels suggests that they resulted from the fusion of a voltage-sensing domain (VSD) to a pore module. Here, we show that the VSD of Ciona intestinalis phosphatase (Ci-VSP) fused to the viral channel Kcv creates Kv(Synth1), a functional voltage-gated, outwardly rectifying K(+) channel. Kv(Synth1) displays the summed features of its individual components: pore properties of Kcv (selectivity and filter gating) and voltage dependence of Ci-VSP (V(1/2) = +56 mV; z of ~1), including the depolarization-induced mode shift. The degree of outward rectification of the channel is critically dependent on the length of the linker more than on its amino acid composition. This highlights a mechanistic role of the linker in transmitting the movement of the sensor to the pore and shows that electromechanical coupling can occur without coevolution of the two domains.
Figures


References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
