

DFMO/eflornithine inhibits migration and invasion downstream of MYCN and involves p27Kip1 activity in neuroblastoma
- PMID: 23440295
- PMCID: PMC3622674
- DOI: 10.3892/ijo.2013.1835
DFMO/eflornithine inhibits migration and invasion downstream of MYCN and involves p27Kip1 activity in neuroblastoma
Abstract
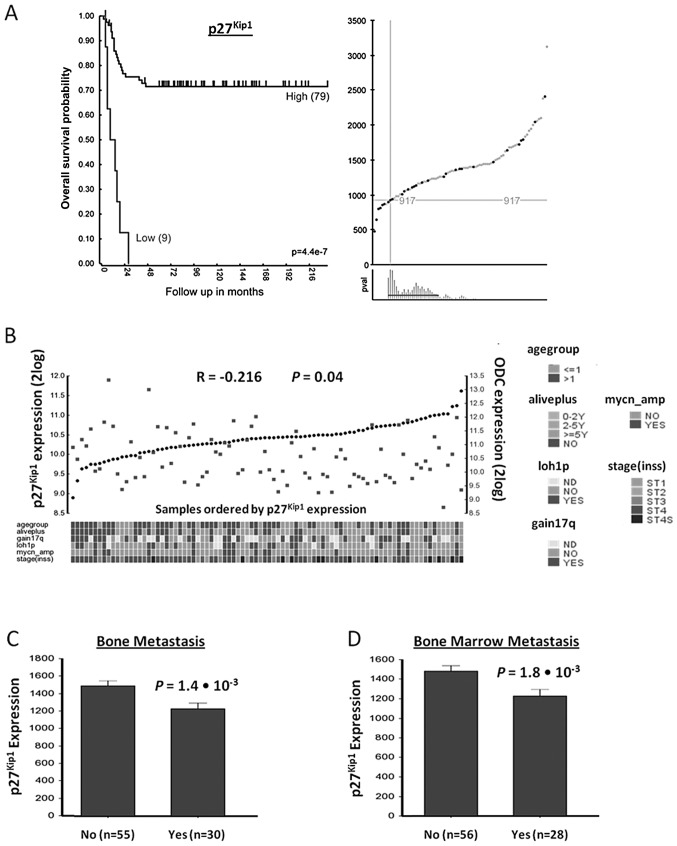
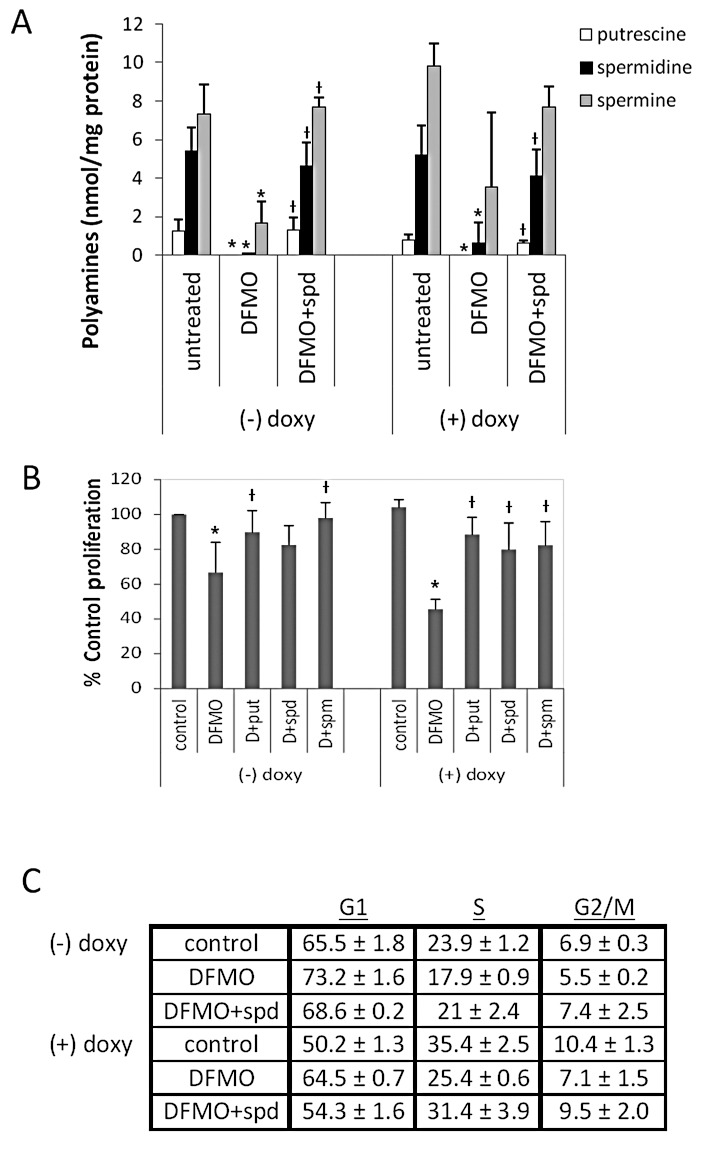
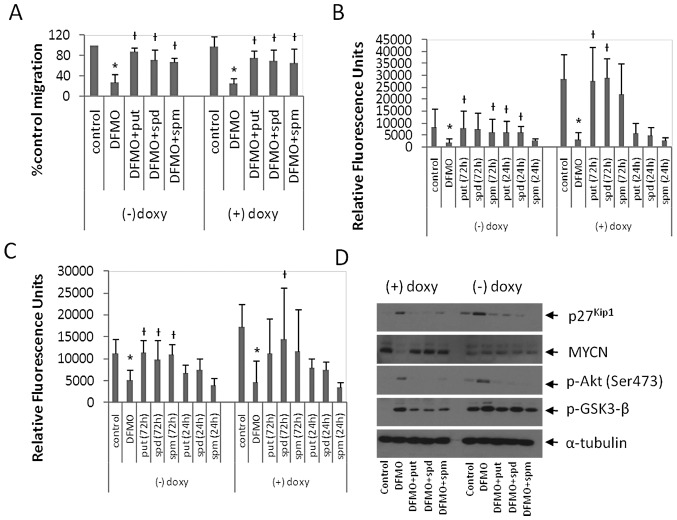
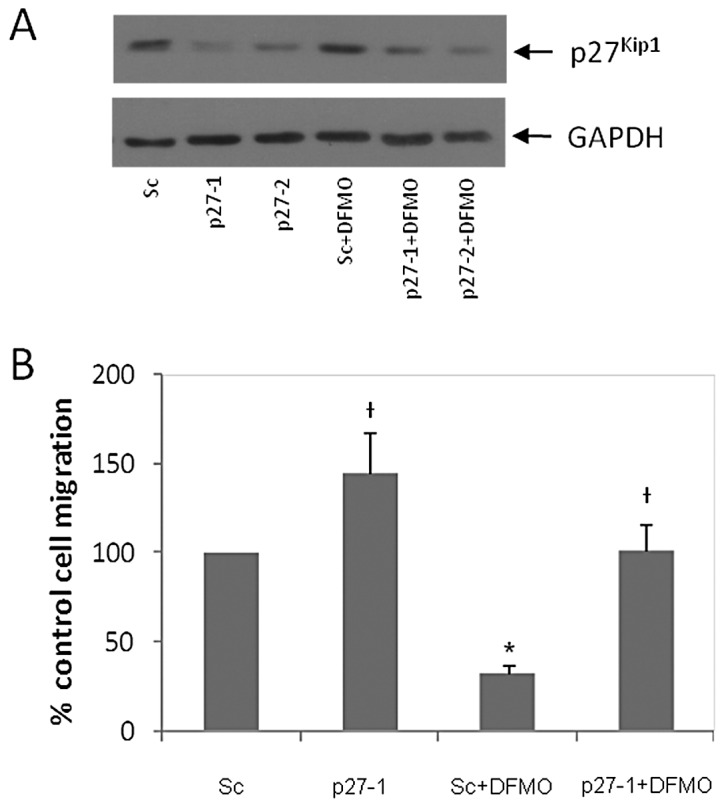
Neuroblastoma (NB) is the most common extracranial pediatric tumor. NB patients over 18 months of age at the time of diagnosis are often in the later stages of the disease, present with widespread dissemination, and often possess MYCN tumor gene amplification. MYCN is a transcription factor that regulates the expression of a number of genes including ornithine decarboxylase (ODC), a rate-limiting enzyme in the biosynthesis of polyamines. Inhibiting ODC in NB cells produces many deleterious effects including G(1) cell cycle arrest, inhibition of cell proliferation, and decreased tumor growth, making ODC a promising target for drug interference. DFMO treatment leads to the accumulation of the cyclin-dependent kinase inhibitor p27(Kip1) protein and causes p27(Kip1)/Rb-coupled G(1) cell cycle arrest in MYCN-amplified NB tumor cells through a process that involves p27(Kip1) phosphorylation at residues Ser10 and Thr198. While p27(Kip1) is well known for its role as a cyclin-dependent kinase inhibitor, recent studies have revealed a novel function of p27(Kip1) as a regulator of cell migration and invasion. In the present study we found that p27(Kip1) regulates the migration and invasion in NB and that these events are dependent on the state of phosphorylation of p27(Kip1). DFMO treatments induced MYCN protein downregulation and phosphorylation of Akt/PKB (Ser473) and GSK3-β (Ser9), and polyamine supplementation alleviated the DFMO-induced effects. Importantly, we provide strong evidence that p27(Kip1) mRNA correlates with clinical features and the survival probability of NB patients.
Figures
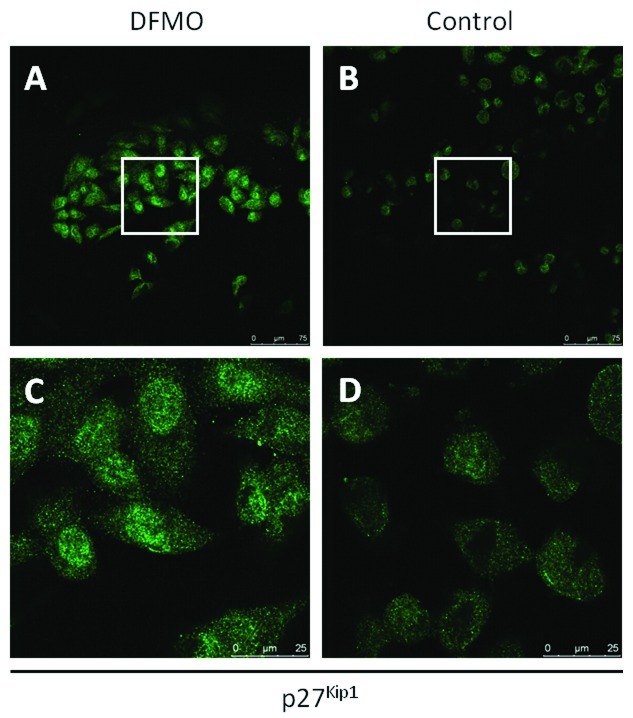
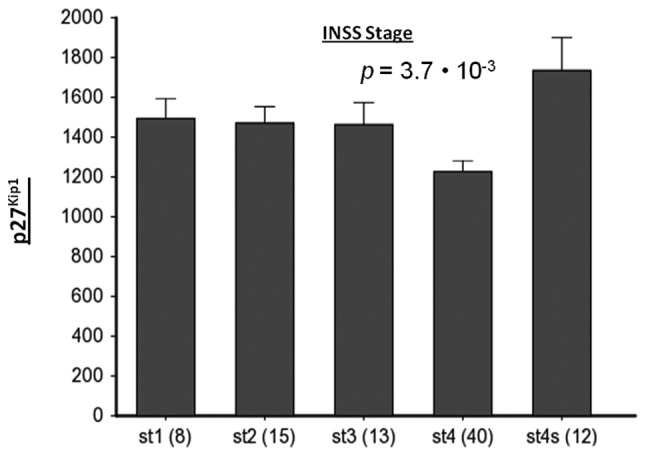
References
-
- Maris JM, Hogarty MD, Bagatell R, Cohn SL. Neuroblastoma. Lancet. 2007;369:2106–2120. - PubMed
-
- Schwab M, Westermann F, Hero B, Berthold F. Neuroblastoma: biology and molecular and chromosomal pathology. Lancet Oncol. 2003;4:472–480. - PubMed
-
- Brodeur GM. Neuroblastoma: biological insights into a clinical enigma. Nat Rev Cancer. 2003;3:203–216. - PubMed
-
- Canete A, Gerrard M, Rubie H, et al. Poor survival for infants with MYCN-amplified metastatic neuroblastoma despite intensified treatment: the International Society of Paediatric Oncology European Neuroblastoma Experience. J Clin Oncol. 2009;27:1014–1019. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
