

Autonomous regulation of the insect gut by circadian genes acting downstream of juvenile hormone signaling
- PMID: 23442387
- PMCID: PMC3600444
- DOI: 10.1073/pnas.1217060110
Autonomous regulation of the insect gut by circadian genes acting downstream of juvenile hormone signaling
Abstract
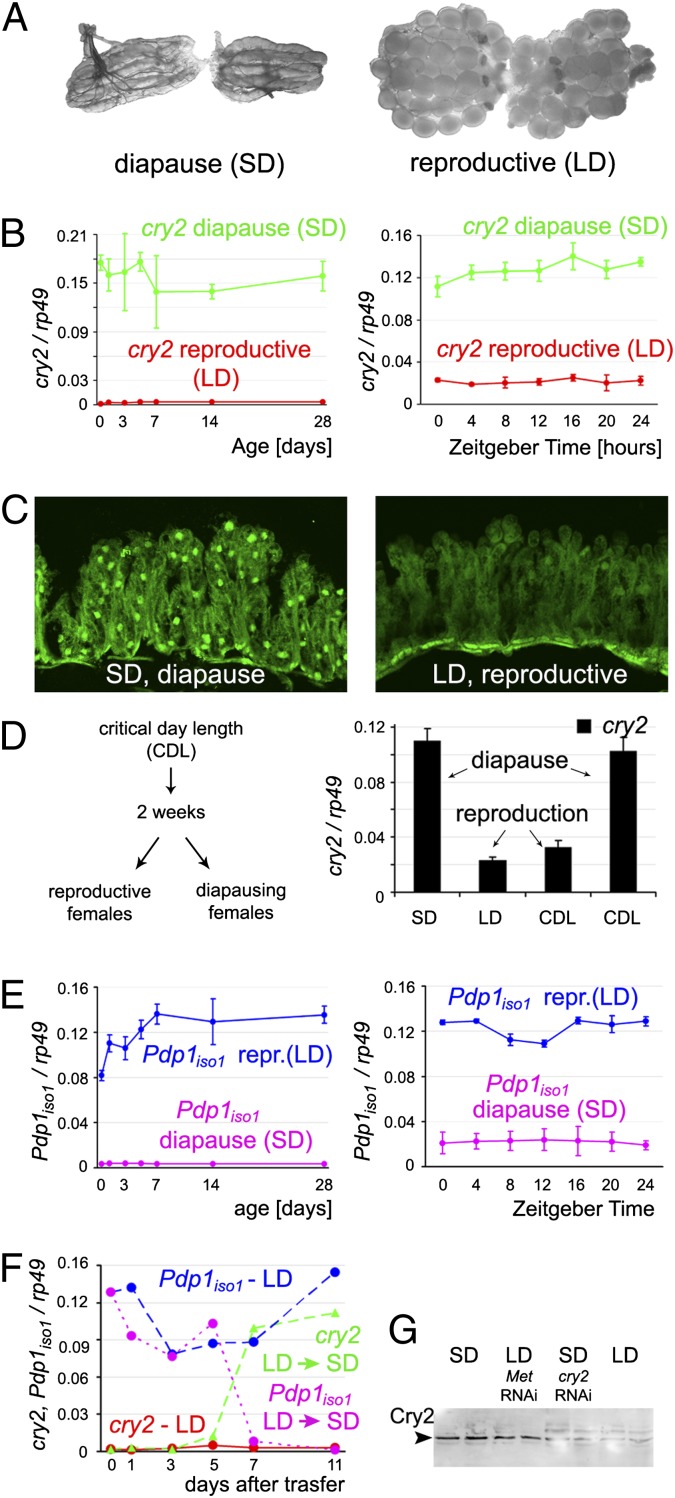
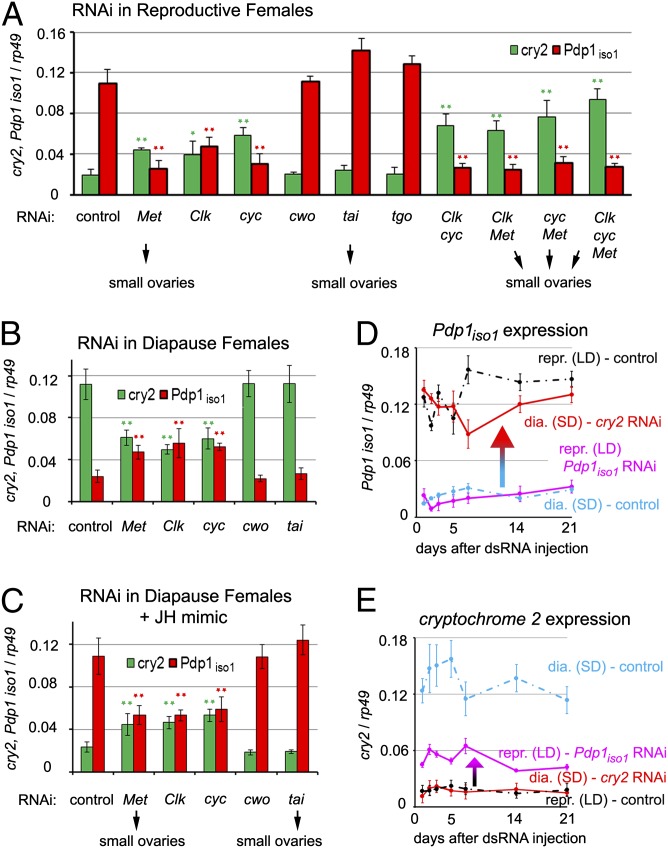
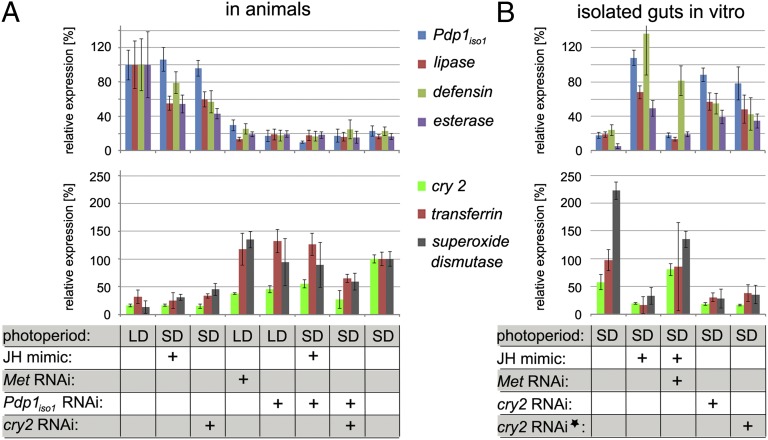
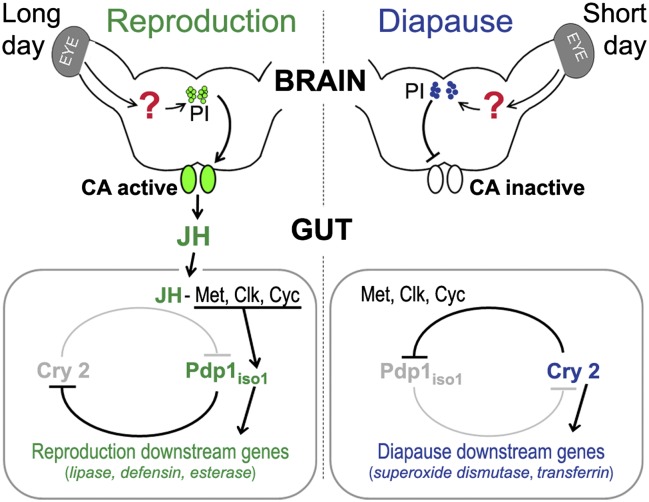
In temperate regions, the shortening day length informs many insect species to prepare for winter by inducing diapause. The adult diapause of the linden bug, Pyrrhocoris apterus, involves a reproductive arrest accompanied by energy storage, reduction of metabolic needs, and preparation to withstand low temperatures. By contrast, nondiapause animals direct nutrient energy to muscle activity and reproduction. The photoperiod-dependent switch from diapause to reproduction is systemically transmitted throughout the organism by juvenile hormone (JH). Here, we show that, at the organ-autonomous level of the insect gut, the decision between reproduction and diapause relies on an interaction between JH signaling and circadian clock genes acting independently of the daily cycle. The JH receptor Methoprene-tolerant and the circadian proteins Clock and Cycle are all required in the gut to activate the Par domain protein 1 gene during reproduction and to simultaneously suppress a mammalian-type cryptochrome 2 gene that promotes the diapause program. A nonperiodic, organ-autonomous feedback between Par domain protein 1 and Cryptochrome 2 then orchestrates expression of downstream genes that mark the diapause vs. reproductive states of the gut. These results show that hormonal signaling through Methoprene-tolerant and circadian proteins controls gut-specific gene activity that is independent of circadian oscillations but differs between reproductive and diapausing animals.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Hahn DA, Denlinger DL. Energetics of insect diapause. Annu Rev Entomol. 2011;56:103–121. - PubMed
-
- Koštál V. Insect photoperiodic calendar and circadian clock: Independence, cooperation, or unity? J Insect Physiol. 2011;57(5):538–556. - PubMed
-
- Bradshaw WE, Holzapfel CM. What season is it anyway? Circadian tracking vs. photoperiodic anticipation in insects. J Biol Rhythms. 2010;25(3):155–165. - PubMed
-
- Hodek I. Diapause in females of Pyrrhocoris apterus L (Heteroptera) Acta Entomol Bohemoslov. 1968;65:422–435.
-
- Slama K. Hormonal control of respiratory metabolism during growth, reproduction, and diapause in female adults of Pyrrhocoris apterus L (Hemiptera) J Insect Physiol. 1964;10:283–303.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
