

Integrating mitochondrial energetics, redox and ROS metabolic networks: a two-compartment model
- PMID: 23442855
- PMCID: PMC3552263
- DOI: 10.1016/j.bpj.2012.11.3808
Integrating mitochondrial energetics, redox and ROS metabolic networks: a two-compartment model
Abstract
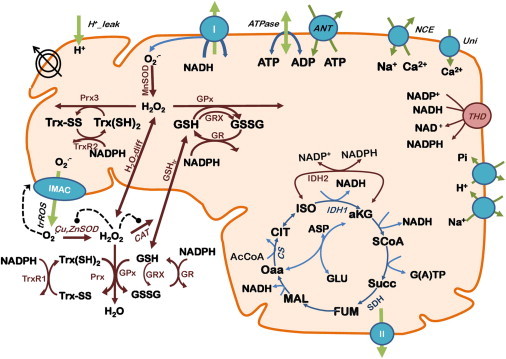
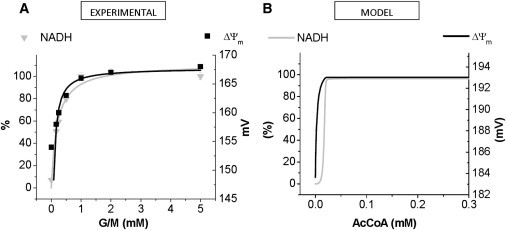
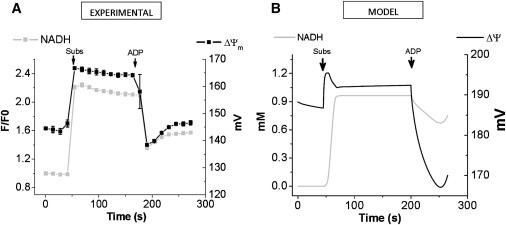
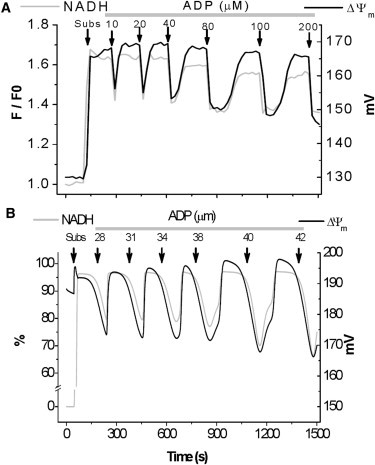
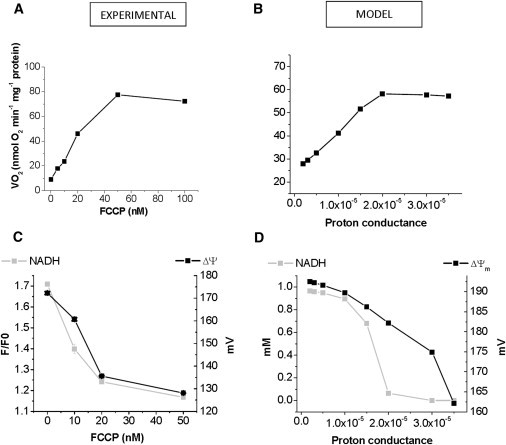
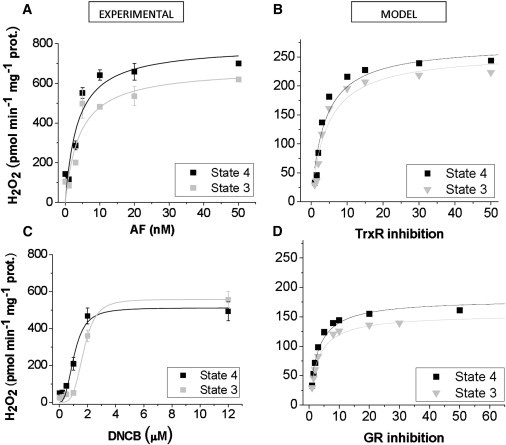
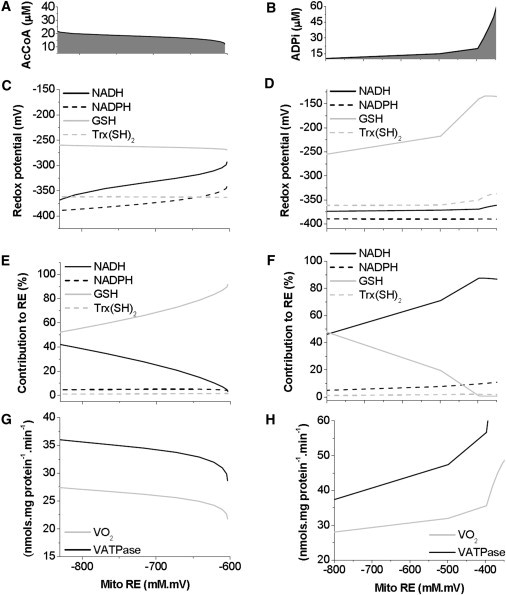
To understand the mechanisms involved in the control and regulation of mitochondrial reactive oxygen species (ROS) levels, a two-compartment computational mitochondrial energetic-redox (ME-R) model accounting for energetic, redox, and ROS metabolisms is presented. The ME-R model incorporates four main redox couples (NADH/NAD(+), NADPH/NADP(+), GSH/GSSG, Trx(SH)(2)/TrxSS). Scavenging systems-glutathione, thioredoxin, superoxide dismutase, catalase-are distributed in mitochondrial matrix and extra-matrix compartments, and transport between compartments of ROS species (superoxide: O(2)(⋅-), hydrogen peroxide: H(2)O(2)), and GSH is also taken into account. Model simulations are compared with experimental data obtained from isolated heart mitochondria. The ME-R model is able to simulate: i), the shape and order of magnitude of H(2)O(2) emission and dose-response kinetics observed after treatment with inhibitors of the GSH or Trx scavenging systems and ii), steady and transient behavior of ΔΨ(m) and NADH after single or repetitive pulses of substrate- or uncoupler-elicited energetic-redox transitions. The dynamics of the redox environment in both compartments is analyzed with the model following substrate addition. The ME-R model represents a useful computational tool for exploring ROS dynamics, the role of compartmentation in the modulation of the redox environment, and how redox regulation participates in the control of mitochondrial function.
Copyright © 2013 Biophysical Society. Published by Elsevier Inc. All rights reserved.
Figures
References
-
- Silverthorn D.U., Ober W.C., Johnson B.R. Pearson Benjamin Cummings; San Francisco, CA: 2009. Human Physiology: An Integrated Approach.
-
- Weibel E.R., Hoppeler H. Exercise-induced maximal metabolic rate scales with muscle aerobic capacity. J. Exp. Biol. 2005;208:1635–1644. - PubMed
-
- Rolfe D.F., Brown G.C. Cellular energy utilization and molecular origin of standard metabolic rate in mammals. Physiol. Rev. 1997;77:731–758. - PubMed
-
- Balaban R.S., Nemoto S., Finkel T. Mitochondria, oxidants, and aging. Cell. 2005;120:483–495. - PubMed
-
- Chance B., Sies H., Boveris A. Hydroperoxide metabolism in mammalian organs. Physiol. Rev. 1979;59:527–605. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
