

Amphipathic lipid packing sensor motifs: probing bilayer defects with hydrophobic residues
- PMID: 23442908
- PMCID: PMC3566459
- DOI: 10.1016/j.bpj.2012.11.3837
Amphipathic lipid packing sensor motifs: probing bilayer defects with hydrophobic residues
Abstract
Sensing membrane curvature allows fine-tuning of complex reactions that occur at the surface of membrane-bound organelles. One of the most sensitive membrane curvature sensors, the Amphipathic Lipid Packing Sensor (ALPS) motif, does not seem to recognize the curved surface geometry of membranes per se; rather, it recognizes defects in lipid packing that arise from membrane bending. In a companion paper, we show that these defects can be mimicked by introducing conical lipids in a flat lipid bilayer, in agreement with experimental observations. Here, we use molecular-dynamics (MD) simulations to characterize ALPS binding to such lipid bilayers. The ALPS motif recognizes lipid-packing defects by a conserved mechanism: peptide partitioning is driven by the insertion of hydrophobic residues into large packing defects that are preformed in the bilayer. This insertion induces only minor modifications in the statistical distribution of the free packing defects. ALPS insertion is severely hampered when monounsaturated lipids are replaced by saturated lipids, leading to a decrease in packing defects. We propose that the hypersensitivity of ALPS motifs to lipid packing defects results from the repetitive use of hydrophobic insertions along the monotonous ALPS sequence.
Copyright © 2013 Biophysical Society. Published by Elsevier Inc. All rights reserved.
Figures
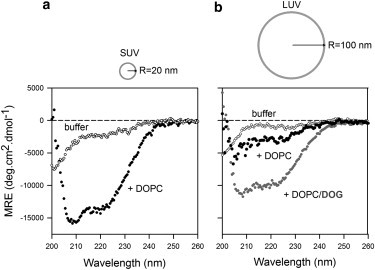

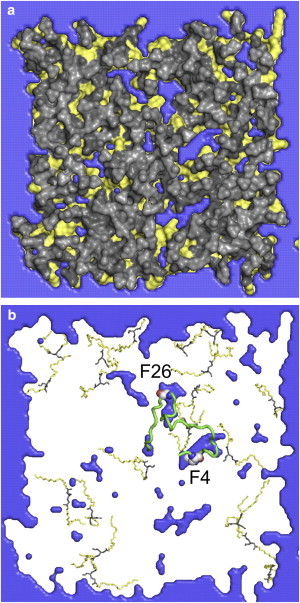

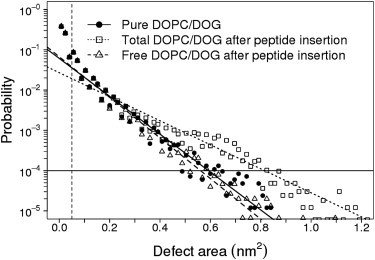


Comment in
-
New and notable: key new insights into membrane targeting by proteins.Biophys J. 2013 Feb 5;104(3):517-9. doi: 10.1016/j.bpj.2012.12.029. Biophys J. 2013. PMID: 23442902 Free PMC article. No abstract available.
References
-
- Antonny B., Huber I., Cassel D. Activation of ADP-ribosylation factor 1 GTPase-activating protein by phosphatidylcholine-derived diacylglycerols. J. Biol. Chem. 1997;272:30848–30851. - PubMed
-
- Bigay J., Gounon P., Antonny B. Lipid packing sensed by ArfGAP1 couples COPI coat disassembly to membrane bilayer curvature. Nature. 2003;426:563–566. - PubMed
-
- McMahon H.T., Gallop J.L. Membrane curvature and mechanisms of dynamic cell membrane remodelling. Nature. 2005;438:590–596. - PubMed
-
- Antonny B. Mechanisms of membrane curvature sensing. Annu. Rev. Biochem. 2011;80:101–123. - PubMed
-
- Zimmerberg J., Kozlov M.M. How proteins produce cellular membrane curvature. Nat. Rev. Mol. Cell Biol. 2006;7:9–19. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
