

Impact of cyanidin-3-glucoside on glycated LDL-induced NADPH oxidase activation, mitochondrial dysfunction and cell viability in cultured vascular endothelial cells
- PMID: 23443099
- PMCID: PMC3546667
- DOI: 10.3390/ijms131215867
Impact of cyanidin-3-glucoside on glycated LDL-induced NADPH oxidase activation, mitochondrial dysfunction and cell viability in cultured vascular endothelial cells
Abstract
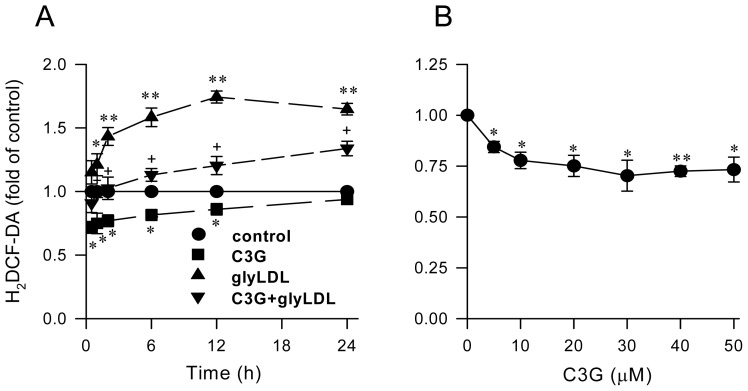
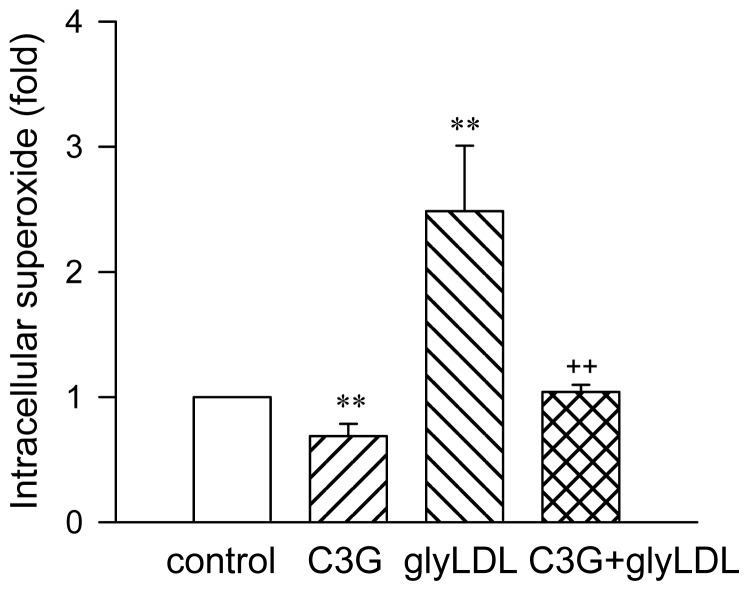
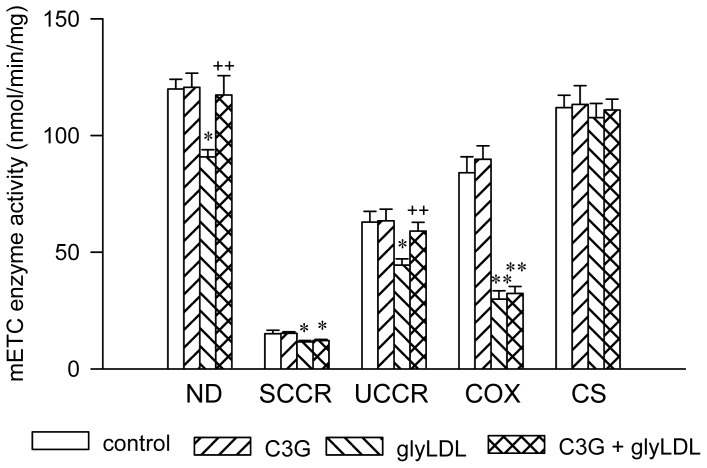
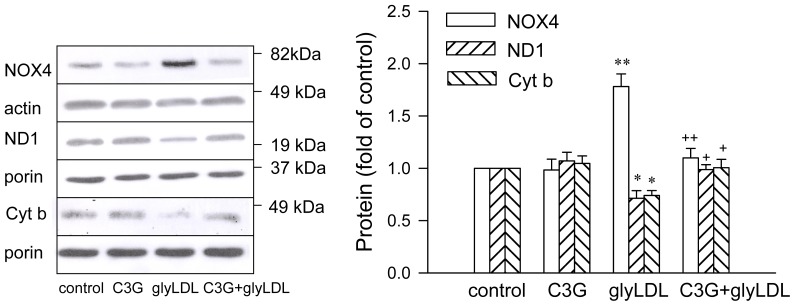
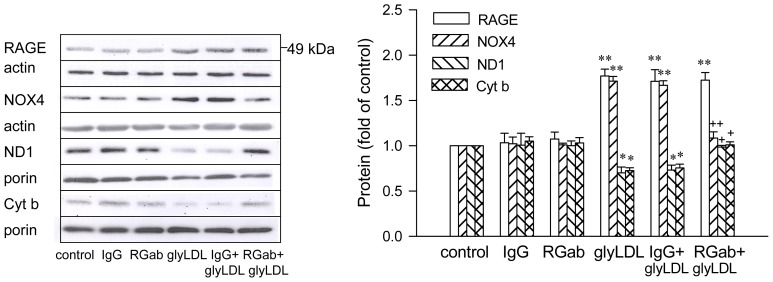
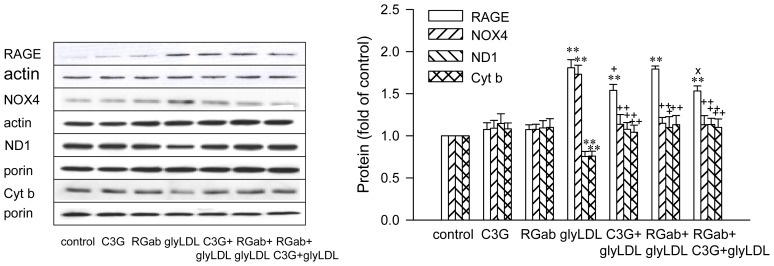
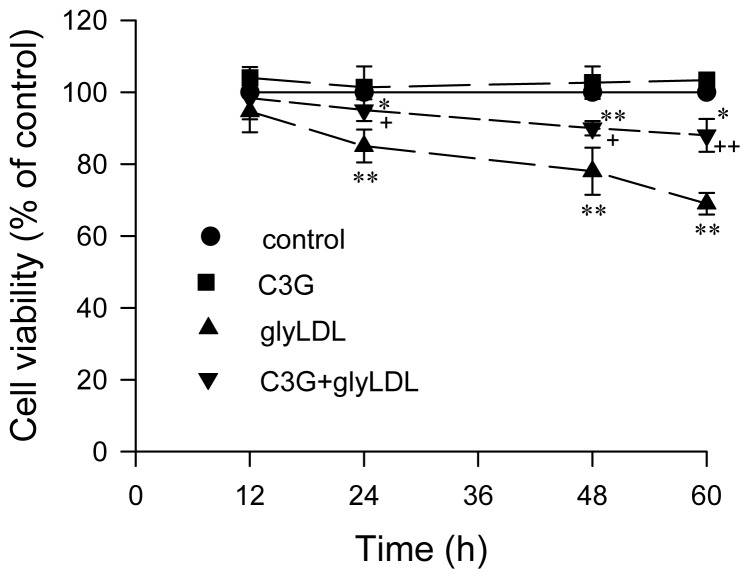
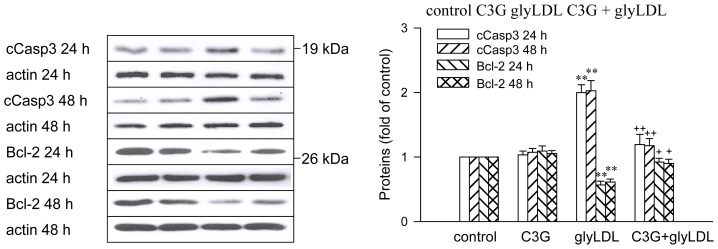
Elevated levels of glycated low density lipoprotein (glyLDL) are frequently detected in diabetic patients. Previous studies demonstrated that glyLDL increased the production of reactive oxygen species (ROS), activated NADPH oxidase (NOX) and suppressed mitochondrial electron transport chain (mETC) enzyme activities in vascular endothelial cells (EC). The present study examined the effects of cyanidin-3-glucoside (C3G), a type of anthocyanin abundant in dark-skinned berries, on glyLDL-induced ROS production, NOX activation and mETC enzyme activity in porcine aortic EC (PAEC). Co-treatment of C3G prevented glyLDL-induced upregulation of NOX4 and intracellular superoxide production in EC. C3G normalized glyLDL-induced inhibition on the enzyme activities of mETC Complex I and III, as well as the abundances of NADH dehydrogenase 1 in Complex I and cytochrome b in Complex III in EC. Blocking antibody for the receptor of advanced glycation end products (RAGE) prevented glyLDL-induced changes in NOX and mETC enzymes. Combination of C3G and RAGE antibody did not significantly enhance glyLDL-induced inhibition of NOX or mETC enzymes. C3G reduced glyLDL-induced RAGE expression with the presence of RAGE antibody. C3G prevented prolonged incubation with the glyLDL-induced decrease in cell viability and the imbalance between key regulators for cell viability (cleaved caspase 3 and B cell Lyphoma-2) in EC. The findings suggest that RAGE plays an important role in glyLDL-induced oxidative stress in vascular EC. C3G may prevent glyLDL-induced NOX activation, the impairment of mETC enzymes and cell viability in cultured vascular EC.
Figures
References
-
- Chao P.C., Huang C.N., Hsu C.C., Yin M.C., Guo Y.R. Association of dietary AGEs with circulating AGEs, glycated LDL, IL-1α and MCP-1 levels in type 2 diabetic patients. Eur. J. Nutr. 2010;49:429–434. - PubMed
-
- Zhao R., Shen G.X. Functional modulation of antioxidant enzymes in vascular endothelial cells by glycated LDL. Atherosclerosis. 2005;179:277–284. - PubMed
-
- Abid M.R., Kachra Z., Spokes K.C., Aird W.C. NADPH oxidase activity is required for endothelial cell proliferation and migration. FEBS Lett. 2000;4862:252–256. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
