

Greater efficiency of photosynthetic carbon fixation due to single amino-acid substitution
- PMID: 23443546
- PMCID: PMC3586729
- DOI: 10.1038/ncomms2504
Greater efficiency of photosynthetic carbon fixation due to single amino-acid substitution
Abstract
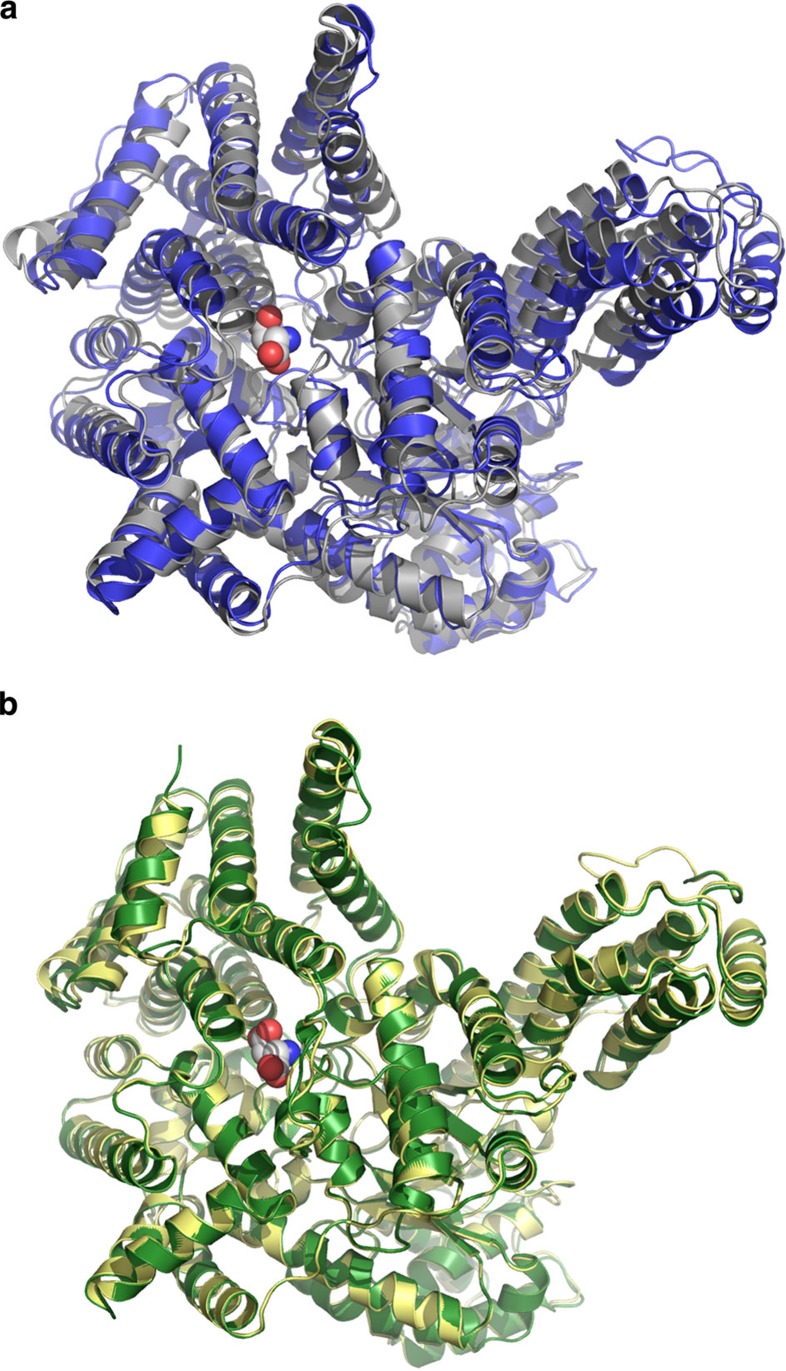
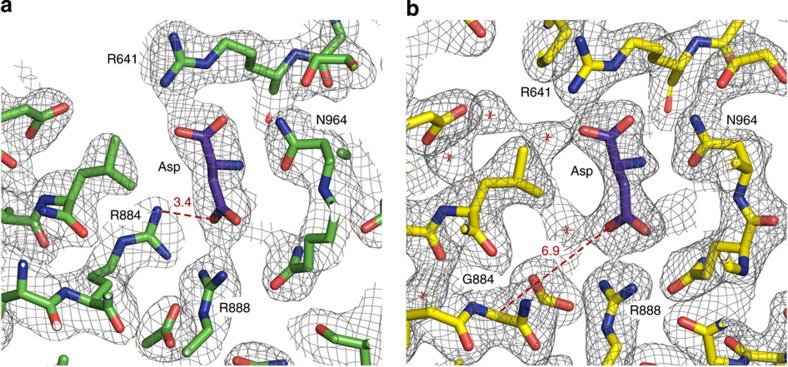
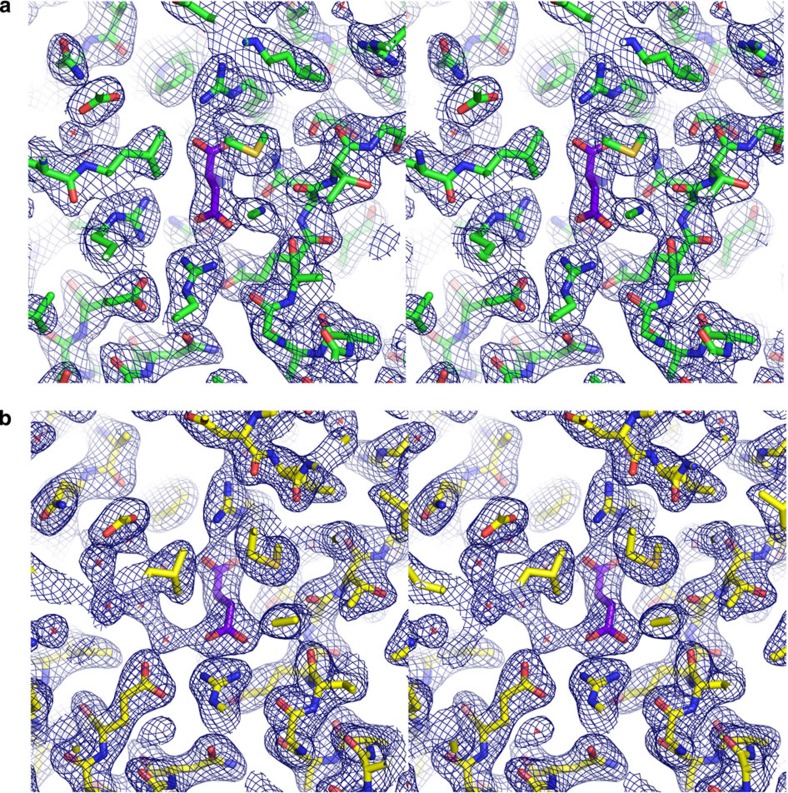
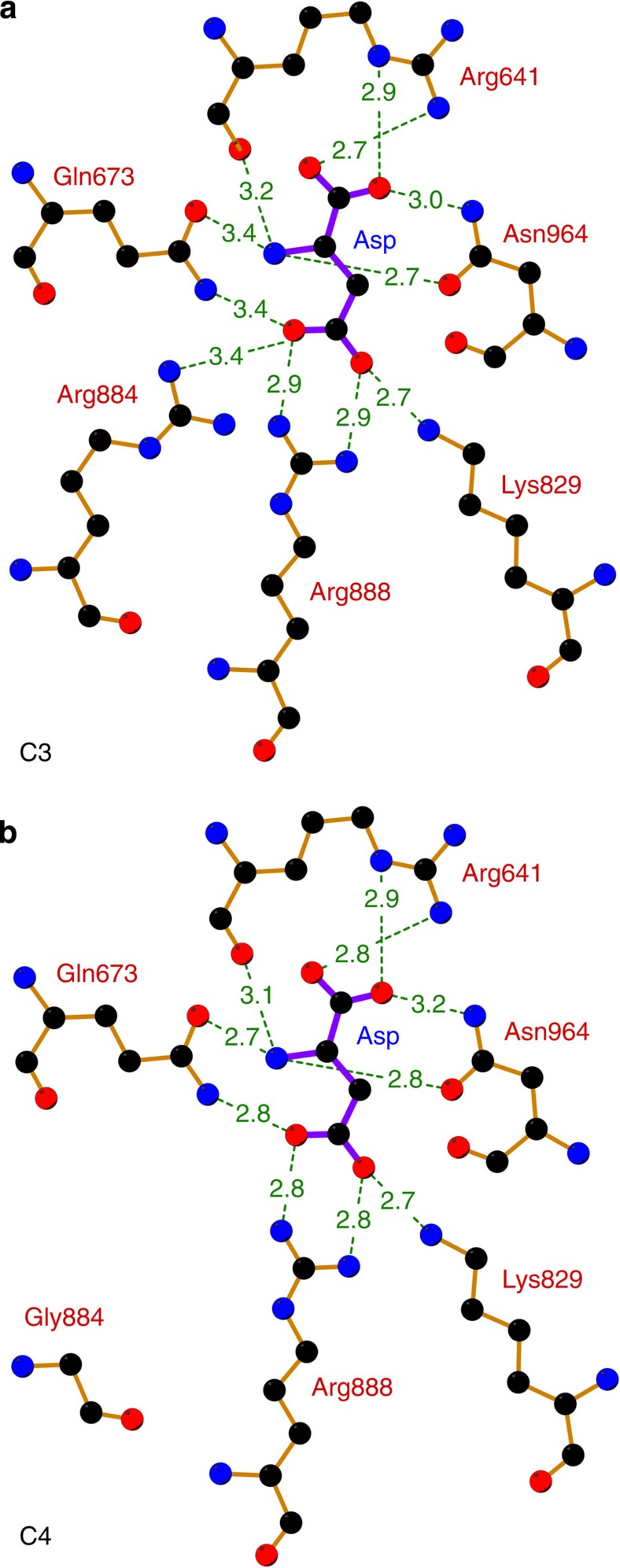
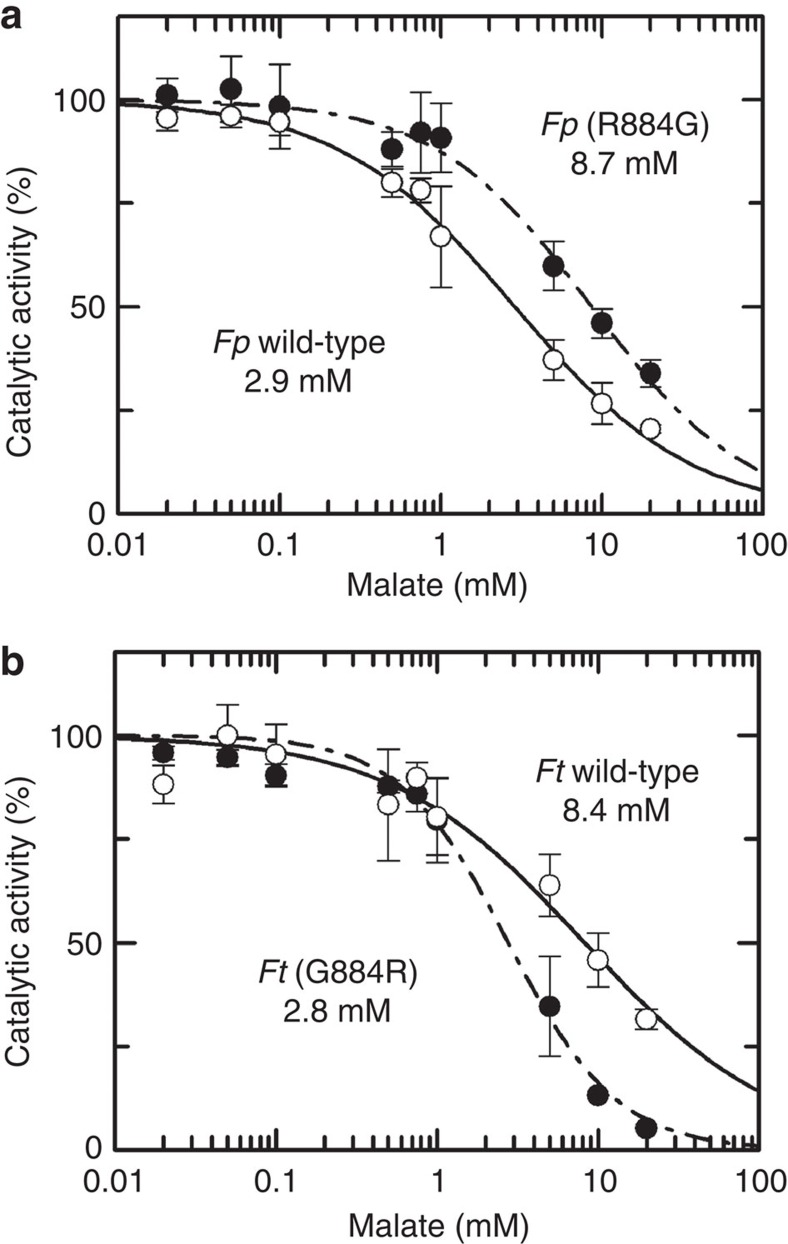
The C4-photosynthetic carbon cycle is an elaborated addition to the classical C3-photosynthetic pathway, which improves solar conversion efficiency. The key enzyme in this pathway, phosphoenolpyruvate carboxylase, has evolved from an ancestral non-photosynthetic C3 phosphoenolpyruvate carboxylase. During evolution, C4 phosphoenolpyruvate carboxylase has increased its kinetic efficiency and reduced its sensitivity towards the feedback inhibitors malate and aspartate. An open question is the molecular basis of the shift in inhibitor tolerance. Here we show that a single-point mutation is sufficient to account for the drastic differences between the inhibitor tolerances of C3 and C4 phosphoenolpyruvate carboxylases. We solved high-resolution X-ray crystal structures of a C3 phosphoenolpyruvate carboxylase and a closely related C4 phosphoenolpyruvate carboxylase. The comparison of both structures revealed that Arg884 supports tight inhibitor binding in the C3-type enzyme. In the C4 phosphoenolpyruvate carboxylase isoform, this arginine is replaced by glycine. The substitution reduces inhibitor affinity and enables the enzyme to participate in the C4 photosynthesis pathway.
Figures

References
-
- Sage R. F.. The evolution of C4 photosynthesis. New Phytol. 161, 341–370 (2004). - PubMed
-
- Sage R. F., Sage T. L. & Kocacinar F.. Photorespiration and the evolution of C4 photosynthesis. Annu. Rev. Plant Physiol. 63, 17.1–17.29 (2012). - PubMed
-
- Stitt M. & Heldt H. W.. Generation and maintenance of concentration gradients between the mesophyll and bundle sheath in maize leaves. Biochim. Biophys. Acta Bioenerg. 808, 400–414 (1985).
-
- Voznesenskaya E. V., Franceschi V. R., Kiirats O., Freitag H. & Edwards G. E.. Kranz anatomy is not essential for terrestrial C4 plant photosynthesis. Nature 414, 543–546 (2001). - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
