

PML bodies provide an important platform for the maintenance of telomeric chromatin integrity in embryonic stem cells
- PMID: 23444137
- PMCID: PMC3632112
- DOI: 10.1093/nar/gkt114
PML bodies provide an important platform for the maintenance of telomeric chromatin integrity in embryonic stem cells
Abstract
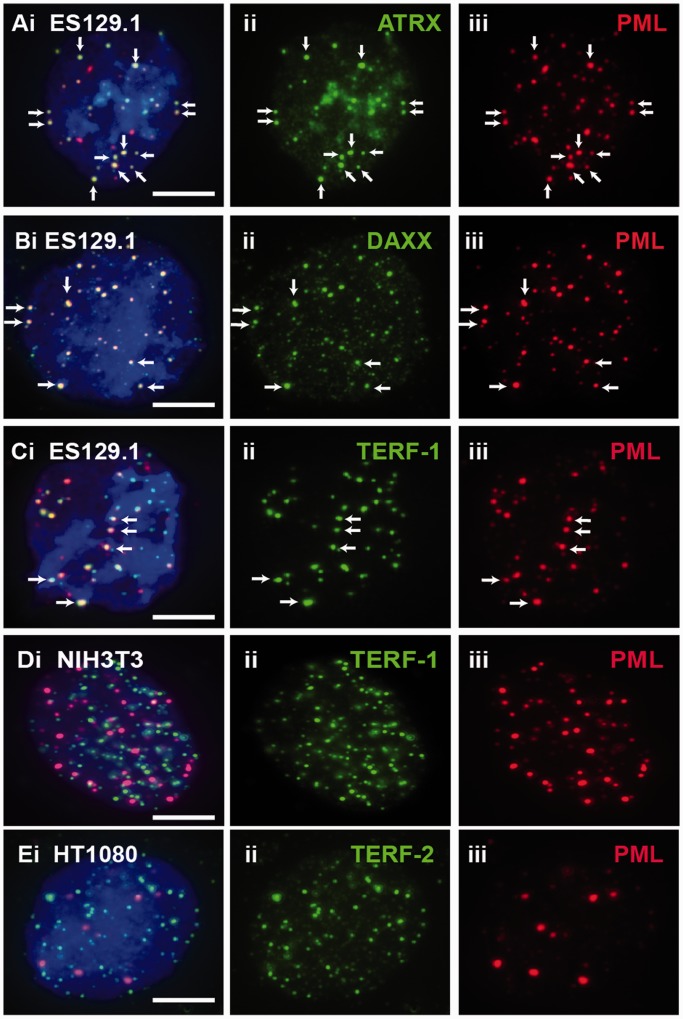
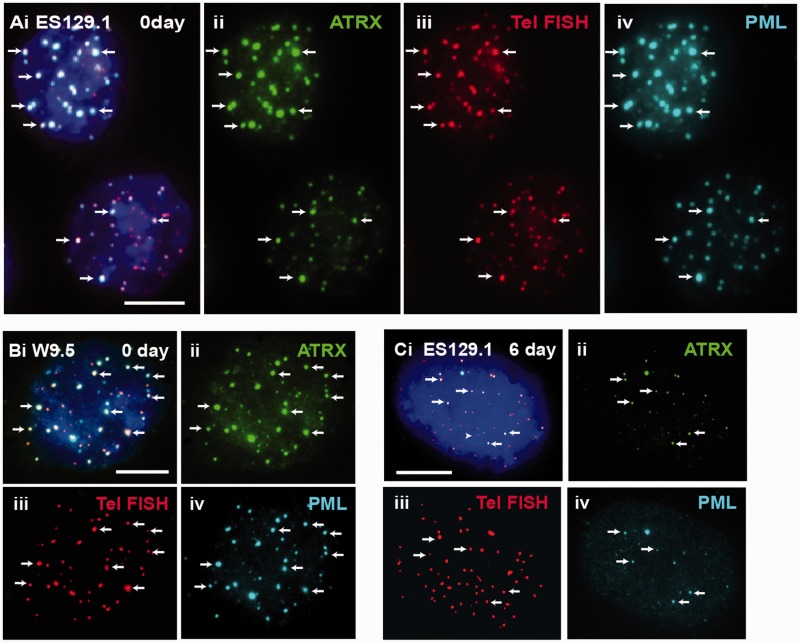
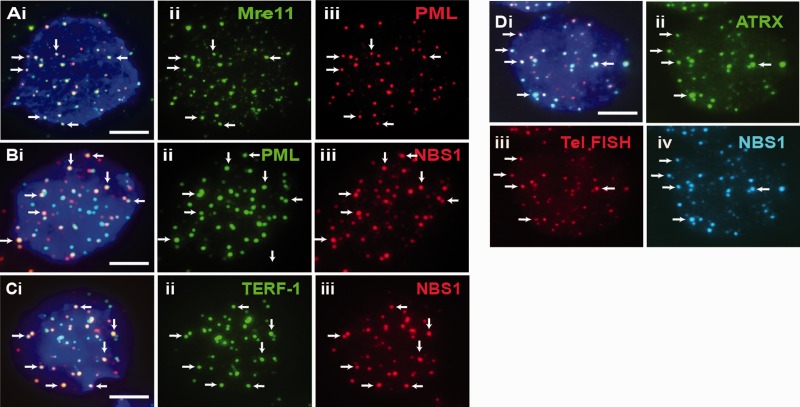
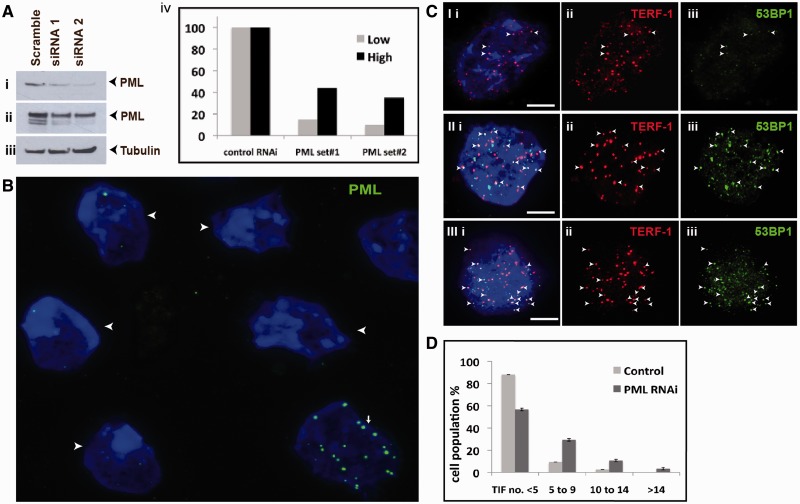
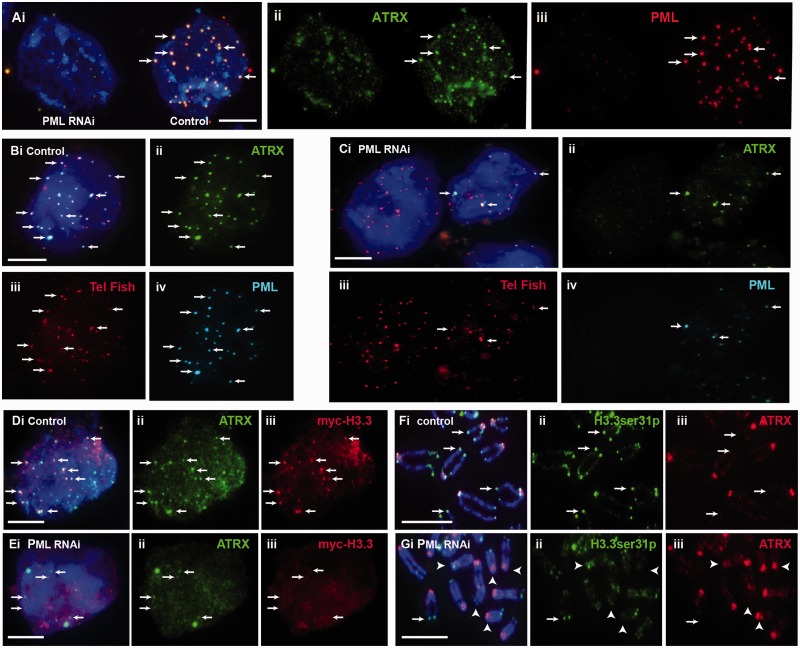
We have previously shown that α-thalassemia mental retardation X-linked (ATRX) and histone H3.3 are key regulators of telomeric chromatin in mouse embryonic stem cells. The function of ATRX and H3.3 in the maintenance of telomere chromatin integrity is further demonstrated by recent studies that show the strong association of ATRX/H3.3 mutations with alternative lengthening of telomeres in telomerase-negative human cancer cells. Here, we demonstrate that ATRX and H3.3 co-localize with the telomeric DNA and associated proteins within the promyelocytic leukemia (PML) bodies in mouse ES cells. The assembly of these telomere-associated PML bodies is most prominent at S phase. RNA interference (RNAi)-mediated knockdown of PML expression induces the disassembly of these nuclear bodies and a telomere dysfunction phenotype in mouse ES cells. Loss of function of PML bodies in mouse ES cells also disrupts binding of ATRX/H3.3 and proper establishment of histone methylation pattern at the telomere. Our study demonstrates that PML bodies act as epigenetic regulators by serving as platforms for the assembly of the telomeric chromatin to ensure a faithful inheritance of epigenetic information at the telomere.
Figures

References
-
- Costa A, Daidone MG, Daprai L, Villa R, Cantu S, Pilotti S, Mariani L, Gronchi A, Henson JD, Reddel RR, et al. Telomere maintenance mechanisms in liposarcomas: association with histologic subtypes and disease progression. Cancer Res. 2006;66:8918–8924. - PubMed
-
- Hakin-Smith V, Jellinek DA, Levy D, Carroll T, Teo M, Timperley WR, McKay MJ, Reddel RR, Royds JA. Alternative lengthening of telomeres and survival in patients with glioblastoma multiforme. Lancet. 2003;361:836–838. - PubMed
-
- Dunham MA, Neumann AA, Fasching CL, Reddel RR. Telomere maintenance by recombination in human cells. Nat. Genet. 2000;26:447–450. - PubMed
-
- Bryan TM, Englezou A, Dalla-Pozza L, Dunham MA, Reddel RR. Evidence for an alternative mechanism for maintaining telomere length in human tumors and tumor-derived cell lines. Nat. Med. 1997;3:1271–1274. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
