

The seirena B class floral homeotic mutant of California Poppy (Eschscholzia californica) reveals a function of the enigmatic PI motif in the formation of specific multimeric MADS domain protein complexes
- PMID: 23444328
- PMCID: PMC3608770
- DOI: 10.1105/tpc.112.105809
The seirena B class floral homeotic mutant of California Poppy (Eschscholzia californica) reveals a function of the enigmatic PI motif in the formation of specific multimeric MADS domain protein complexes
Abstract
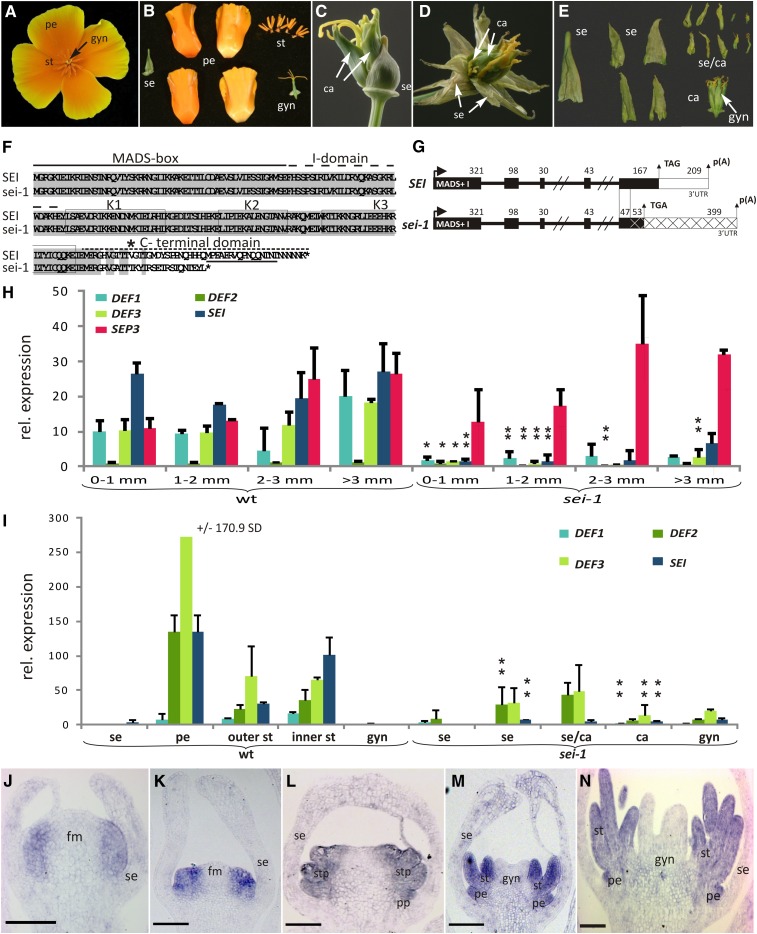
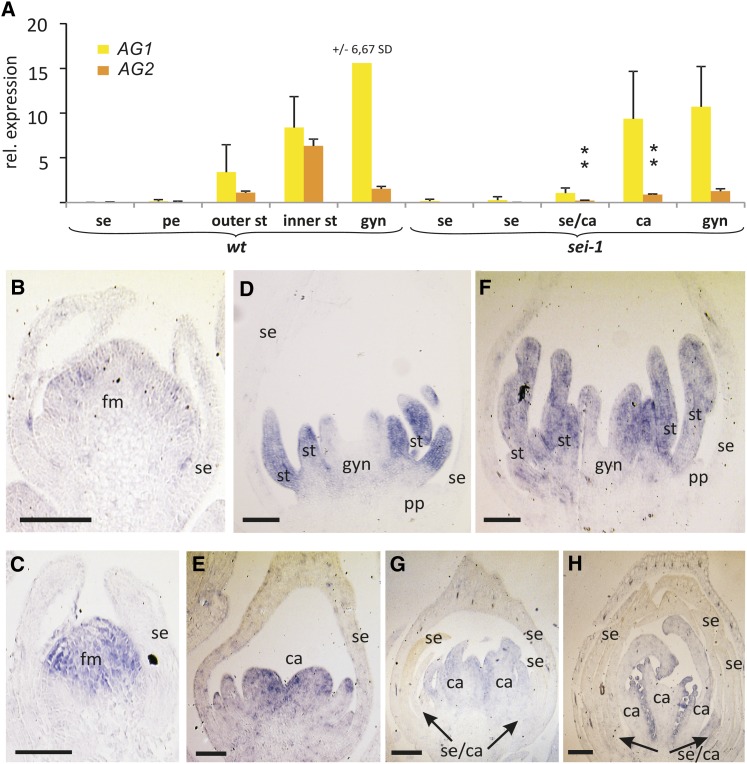
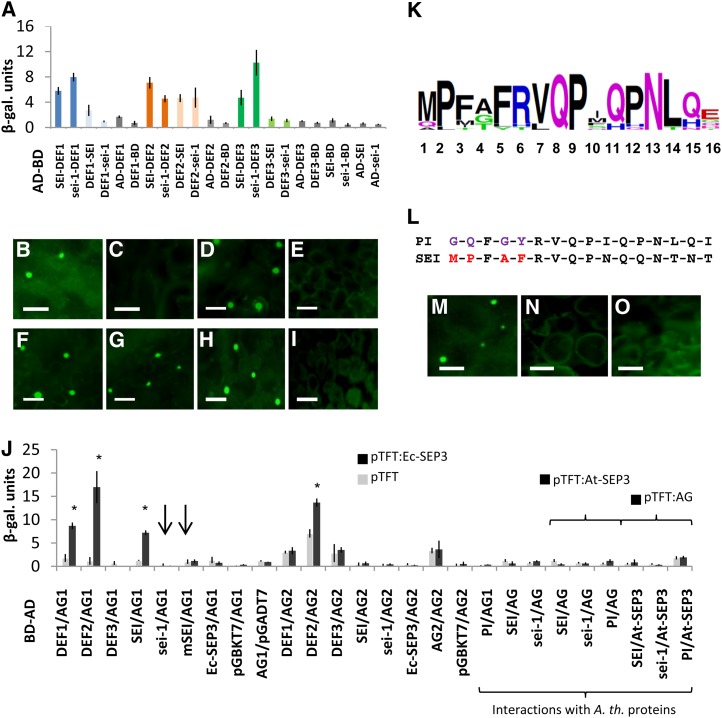
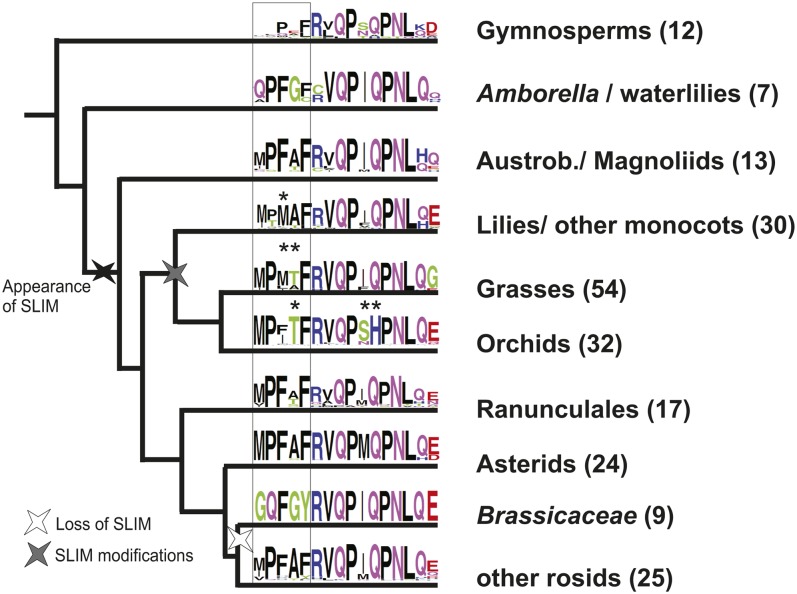
The products of B class floral homeotic genes specify petal and stamen identity, and loss of B function results in homeotic conversions of petals into sepals and stamens into carpels. Here, we describe the molecular characterization of seirena-1 (sei-1), a mutant from the basal eudicot California poppy (Eschscholzia californica) that shows homeotic changes characteristic of floral homeotic B class mutants. SEI has been previously described as EScaGLO, one of four B class-related MADS box genes in California poppy. The C terminus of SEI, including the highly conserved PI motif, is truncated in sei-1 proteins. Nevertheless, like the wild-type SEI protein, the sei-1 mutant protein is able to bind CArG-boxes and can form homodimers, heterodimers, and several higher order complexes with other MADS domain proteins. However, unlike the wild type, the mutant protein is not able to mediate higher order complexes consisting of specific B, C, and putative E class related proteins likely involved in specifying stamen identity. Within the PI motif, five highly conserved N-terminal amino acids are specifically required for this interaction. Several families lack this short conserved sequence, including the Brassicaceae, and we propose an evolutionary scenario to explain these functional differences.
Figures
Similar articles
-
Floral homeotic C function genes repress specific B function genes in the carpel whorl of the basal eudicot California poppy (Eschscholzia californica).Evodevo. 2010 Dec 1;1:13. doi: 10.1186/2041-9139-1-13. Evodevo. 2010. PMID: 21122096 Free PMC article.
-
The study of the E-class SEPALLATA3-like MADS-box genes in wild-type and mutant flowers of cultivated saffron crocus (Crocus sativus L.) and its putative progenitors.J Plant Physiol. 2011 Sep 15;168(14):1675-84. doi: 10.1016/j.jplph.2011.03.015. Epub 2011 May 31. J Plant Physiol. 2011. PMID: 21621873
-
Comparative transcriptomics among floral organs of the basal eudicot Eschscholzia californica as reference for floral evolutionary developmental studies.Genome Biol. 2010;11(10):R101. doi: 10.1186/gb-2010-11-10-r101. Epub 2010 Oct 15. Genome Biol. 2010. PMID: 20950453 Free PMC article.
-
Tinkering with transcription factor networks for developmental robustness of Ranunculales flowers.Ann Bot. 2016 Apr;117(5):845-58. doi: 10.1093/aob/mcw037. Epub 2016 Apr 18. Ann Bot. 2016. PMID: 27091506 Free PMC article. Review.
-
The 'ABC' of MADS domain protein behaviour and interactions.Semin Cell Dev Biol. 2010 Feb;21(1):87-93. doi: 10.1016/j.semcdb.2009.10.004. Epub 2009 Oct 31. Semin Cell Dev Biol. 2010. PMID: 19883778 Review.
Cited by
-
CsPI from the perianthless early-diverging Chloranthus spicatus show function on petal development in Arabidopsis thaliana.Bot Stud. 2014 Dec;55(1):21. doi: 10.1186/1999-3110-55-21. Epub 2014 Feb 4. Bot Stud. 2014. PMID: 28510925 Free PMC article.
-
Arabidopsis flower development--of protein complexes, targets, and transport.Protoplasma. 2016 Mar;253(2):219-30. doi: 10.1007/s00709-015-0812-7. Epub 2015 Apr 7. Protoplasma. 2016. PMID: 25845756 Review.
-
The Maize PI/GLO Ortholog Zmm16/sterile tassel silky ear1 Interacts with the Zygomorphy and Sex Determination Pathways in Flower Development.Plant Cell. 2015 Nov;27(11):3081-98. doi: 10.1105/tpc.15.00679. Epub 2015 Oct 30. Plant Cell. 2015. PMID: 26518212 Free PMC article.
-
Deciphering the Physalis floridana double-layered-lantern1 mutant provides insights into functional divergence of the GLOBOSA duplicates within the Solanaceae.Plant Physiol. 2014 Feb;164(2):748-64. doi: 10.1104/pp.113.233072. Epub 2014 Jan 3. Plant Physiol. 2014. PMID: 24390390 Free PMC article.
-
A cornucopia of diversity-Ranunculales as a model lineage.J Exp Bot. 2024 Mar 27;75(7):1800-1822. doi: 10.1093/jxb/erad492. J Exp Bot. 2024. PMID: 38109712 Free PMC article. Review.
References
-
- Ambrose B.A., Lerner D.R., Ciceri P., Padilla C.M., Yanofsky M.F., Schmidt R.J. (2000). Molecular and genetic analyses of the silky1 gene reveal conservation in floral organ specification between eudicots and monocots. Mol. Cell 5: 569–579 - PubMed
-
- Becker A., Gleissberg S., Smyth D.R. (2005). Floral and vegetative morphogenesis in California poppy (Eschscholzia californica Cham.). Int. J. Plant Sci. 166: 537–555
-
- Benlloch R., Roque E., Ferrándiz C., Cosson V., Caballero T., Penmetsa R.V., Beltrán J.P., Cañas L.A., Ratet P., Madueño F. (2009). Analysis of B function in legumes: PISTILLATA proteins do not require the PI motif for floral organ development in Medicago truncatula. Plant J. 60: 102–111 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
