

Neonatal systemic exposure to lipopolysaccharide enhances susceptibility of nigrostriatal dopaminergic neurons to rotenone neurotoxicity in later life
- PMID: 23446007
- PMCID: PMC3777222
- DOI: 10.1159/000346156
Neonatal systemic exposure to lipopolysaccharide enhances susceptibility of nigrostriatal dopaminergic neurons to rotenone neurotoxicity in later life
Abstract
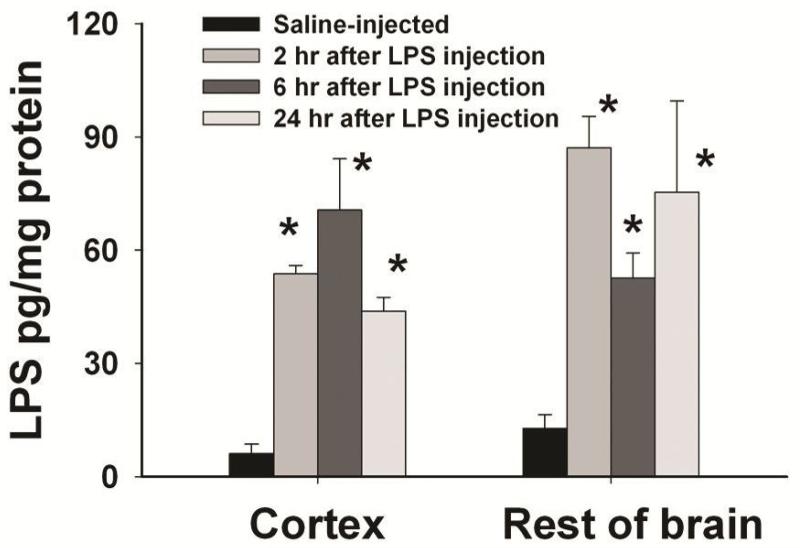
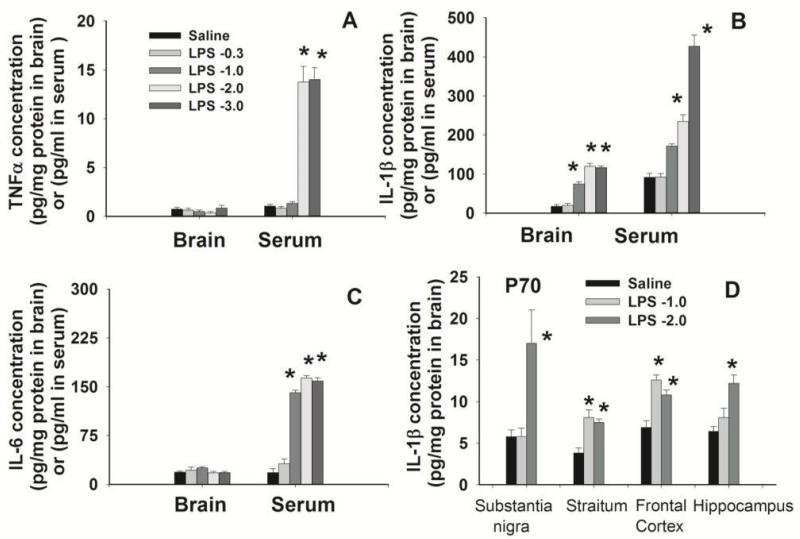
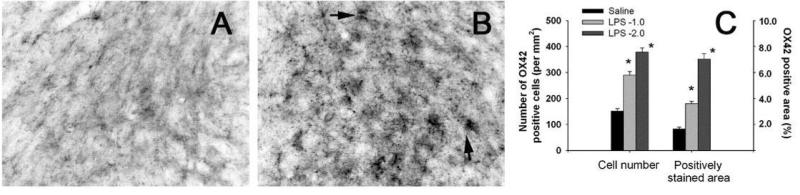
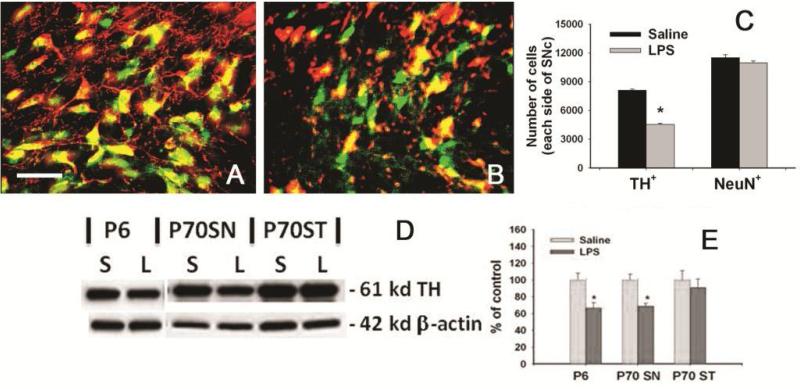
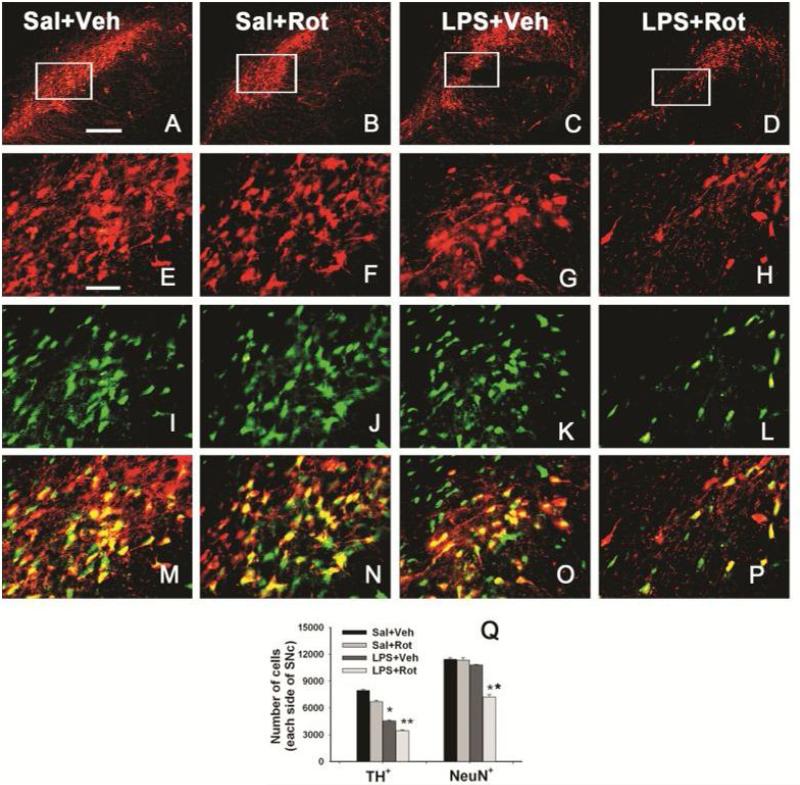
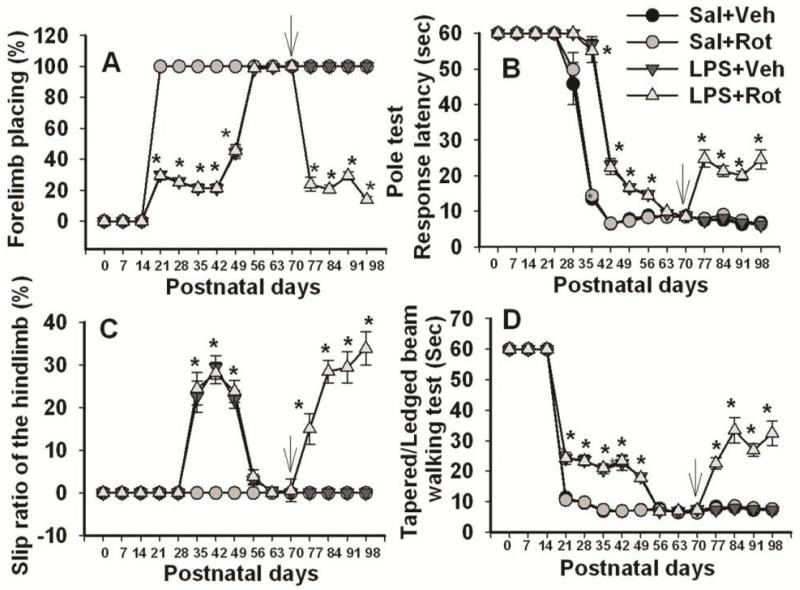
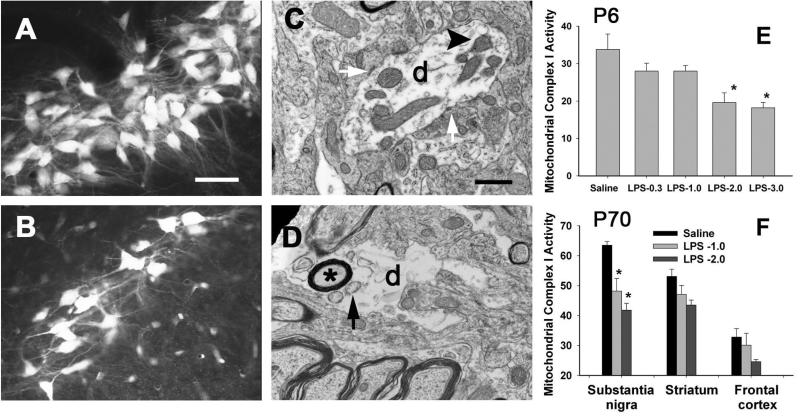
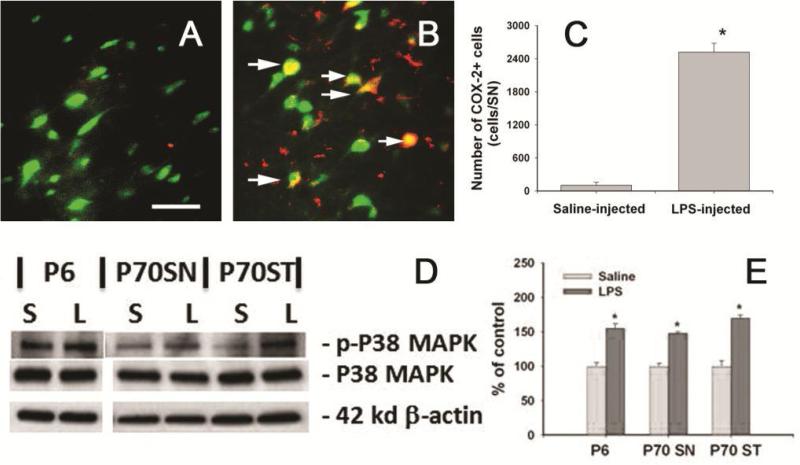
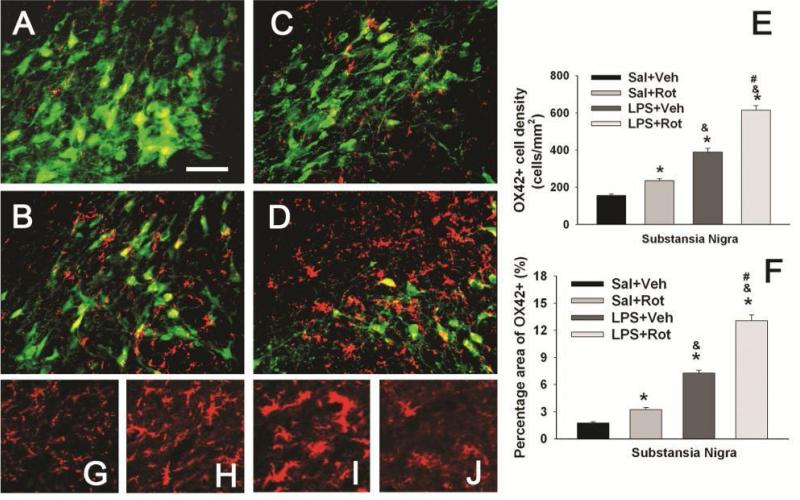
Brain inflammation via intracerebral injection with lipopolysaccharide (LPS) in early life has been shown to increase risks for the development of neurodegenerative disorders in adult rats. To determine if neonatal systemic LPS exposure has the same effects on enhancement of adult dopaminergic neuron susceptibility to rotenone neurotoxicity as centrally injected LPS does, LPS (2 μg/g body weight) was administered intraperitoneally into postnatal day 5 (P5) rats and when grown to P70, rats were challenged with rotenone, a commonly used pesticide, through subcutaneous minipump infusion at a dose of 1.25 mg/kg/day for 14 days. Systemically administered LPS can penetrate into the neonatal rat brain and cause acute and chronic brain inflammation, as evidenced by persistent increases in IL-1β levels, cyclooxygenase-2 expression and microglial activation in the substantia nigra (SN) of P70 rats. Neonatal LPS exposure resulted in suppression of tyrosine hydroxylase (TH) expression, but not actual death of dopaminergic neurons in the SN, as indicated by the reduced number of TH+ cells and unchanged total number of neurons (NeuN+) in the SN. Neonatal LPS exposure also caused motor function deficits, which were spontaneously recoverable by P70. A small dose of rotenone at P70 induced loss of dopaminergic neurons, as indicated by reduced numbers of both TH+ and NeuN+ cells in the SN, and Parkinson's disease (PD)-like motor impairment in P98 rats that had experienced neonatal LPS exposure, but not in those without the LPS exposure. These results indicate that although neonatal systemic LPS exposure may not necessarily lead to death of dopaminergic neurons in the SN, such an exposure could cause persistent functional alterations in the dopaminergic system and indirectly predispose the nigrostriatal system in the adult brain to be damaged by environmental toxins at an ordinarily nontoxic or subtoxic dose and develop PD-like pathological features and motor dysfunction.
Copyright © 2013 S. Karger AG, Basel.
Figures
References
-
- Hagberg H, Gressens P, Mallard C. Inflammation during fetal and neonatal life: implications for neurologic and neuropsychiatric disease in children and adults. Ann Neurol. 2012;71:444–457. - PubMed
-
- Meyer U, Yee BK, Feldon J. The neurodevelopmental impact of prenatal infections at different times of pregnancy: the earlier the worse? Neuroscientist. 2007;13:241–256. - PubMed
-
- Meyer U, Feldon J, Fatemi SH. In-vivo rodent models for the experimental investigation of prenatal immune activation effects in neurodevelopmental brain disorders. Neurosci Biobehav Rev. 2009;33:1061–1079. - PubMed
-
- Patterson PH. Neuroscience. Maternal effects on Schizophrenia risk. Science. 2007;318:576–577. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
