

Animal-to-animal variability in the phasing of the crustacean cardiac motor pattern: an experimental and computational analysis
- PMID: 23446690
- PMCID: PMC3653045
- DOI: 10.1152/jn.01010.2012
Animal-to-animal variability in the phasing of the crustacean cardiac motor pattern: an experimental and computational analysis
Erratum in
- J Neurophysiol. 2013 Aug;110(4):1036
Abstract
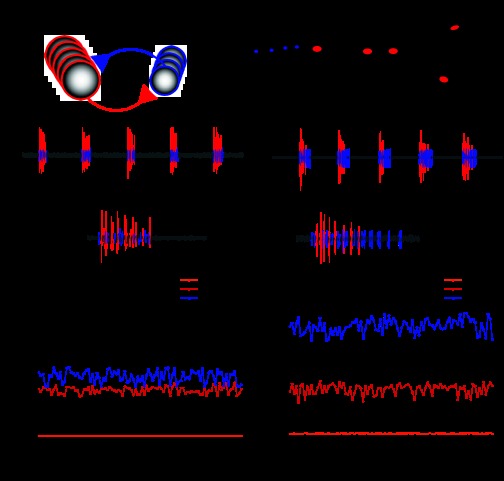
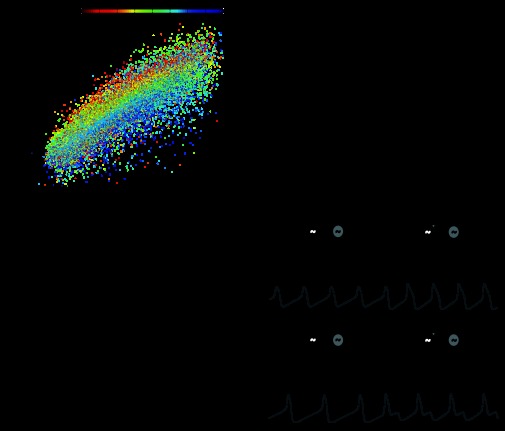
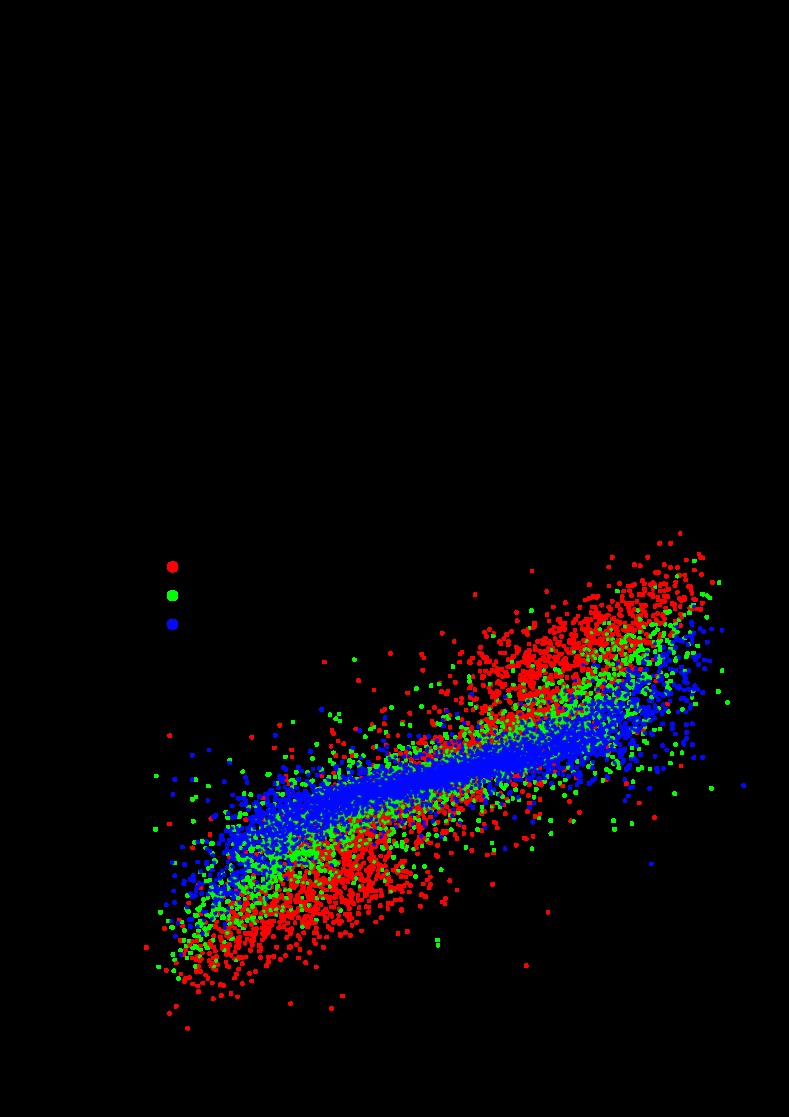
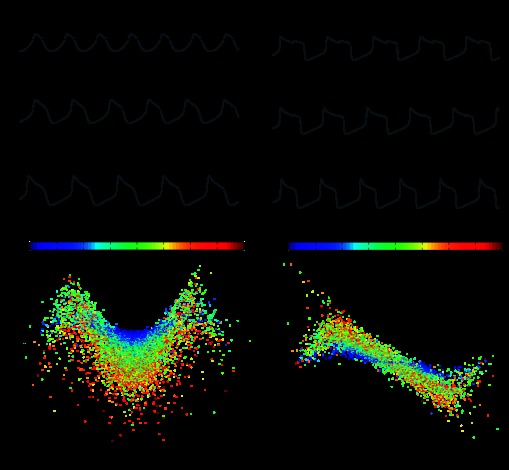
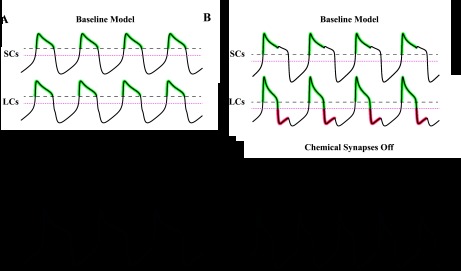
The cardiac ganglion (CG) of Homarus americanus is a central pattern generator that consists of two oscillatory groups of neurons: "small cells" (SCs) and "large cells" (LCs). We have shown that SCs and LCs begin their bursts nearly simultaneously but end their bursts at variable phases. This variability contrasts with many other central pattern generator systems in which phase is well maintained. To determine both the consequences of this variability and how CG phasing is controlled, we modeled the CG as a pair of Morris-Lecar oscillators coupled by electrical and excitatory synapses and constructed a database of 15,000 simulated networks using random parameter sets. These simulations, like our experimental results, displayed variable phase relationships, with the bursts beginning together but ending at variable phases. The model suggests that the variable phasing of the pattern has important implications for the functional role of the excitatory synapses. In networks in which the two oscillators had similar duty cycles, the excitatory coupling functioned to increase cycle frequency. In networks with disparate duty cycles, it functioned to decrease network frequency. Overall, we suggest that the phasing of the CG may vary without compromising appropriate motor output and that this variability may critically determine how the network behaves in response to manipulations.
Keywords: central pattern generator; morris-lecar model; neuronal oscillators; phase relationships.
Figures




References
-
- Berens P. Journal of Statistical Software. CircStat: a Matlab Toolbox for Circular Statistics (online). http://www.jstatsoft.org/v31/i10 [7 March 2013].
-
- Berlind A. Heterogeneity of motorneuron driver potential properties along the anterior-posterior axis of the lobster cardiac ganglion. Brain Res 609: 51–58, 1993 - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources