

Increased agonist affinity at the μ-opioid receptor induced by prolonged agonist exposure
- PMID: 23447620
- PMCID: PMC3711766
- DOI: 10.1523/JNEUROSCI.4187-12.2013
Increased agonist affinity at the μ-opioid receptor induced by prolonged agonist exposure
Abstract
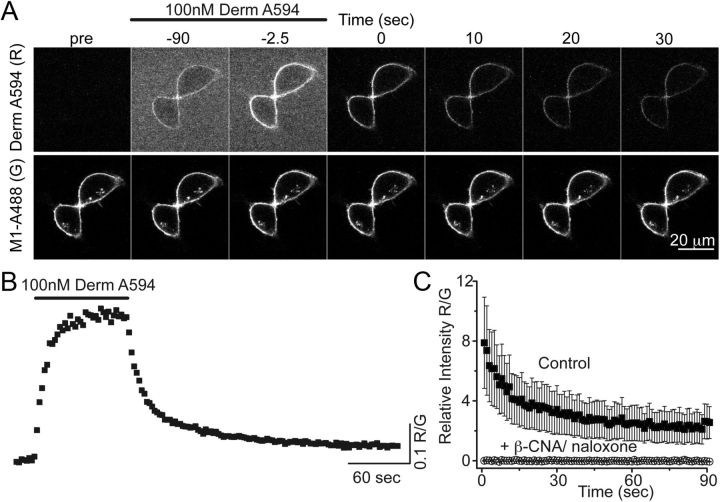
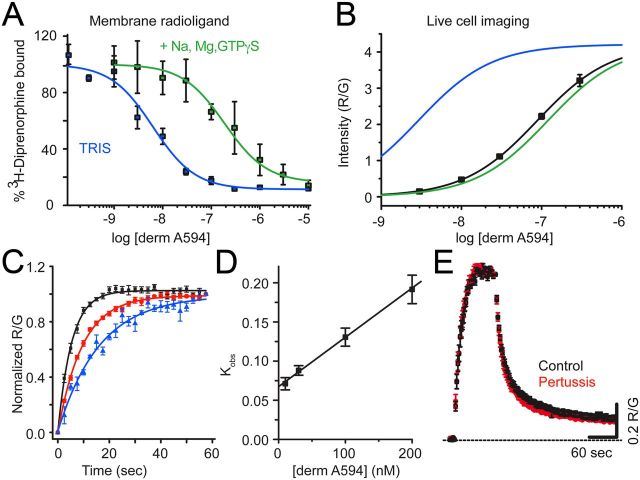
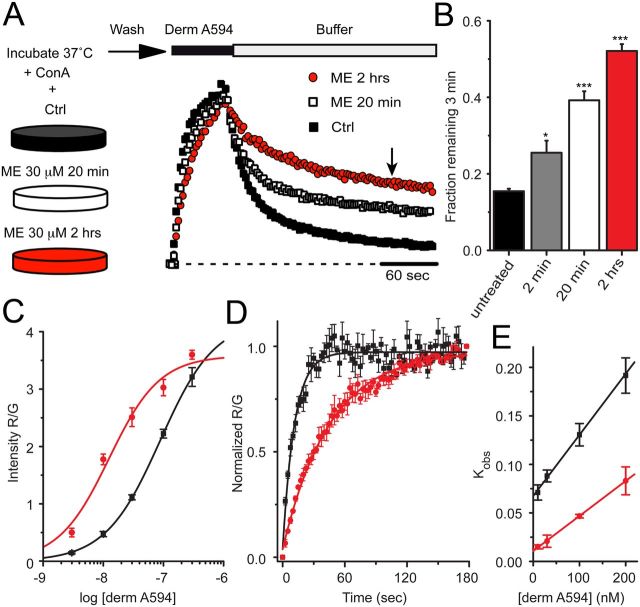
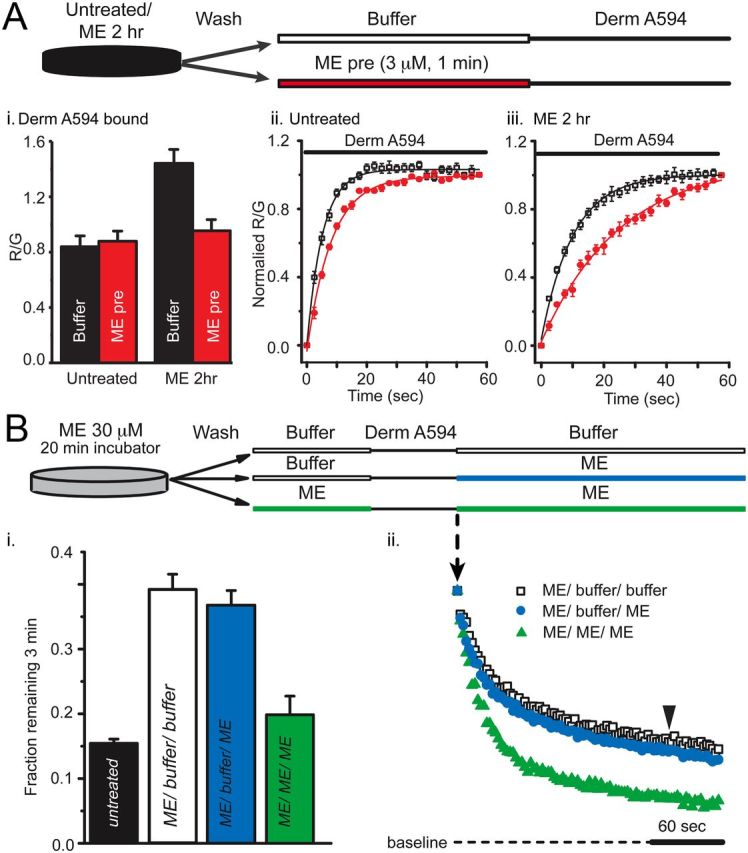
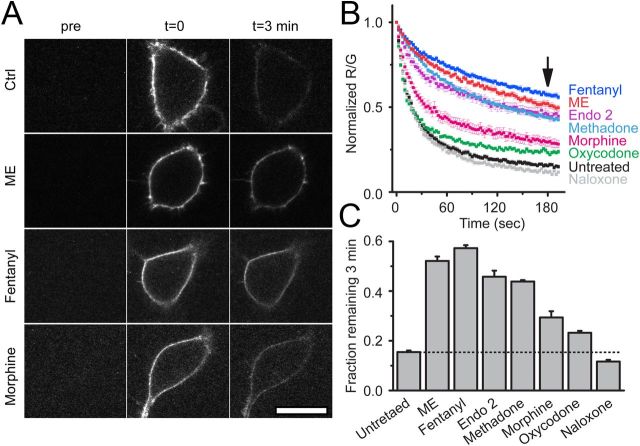
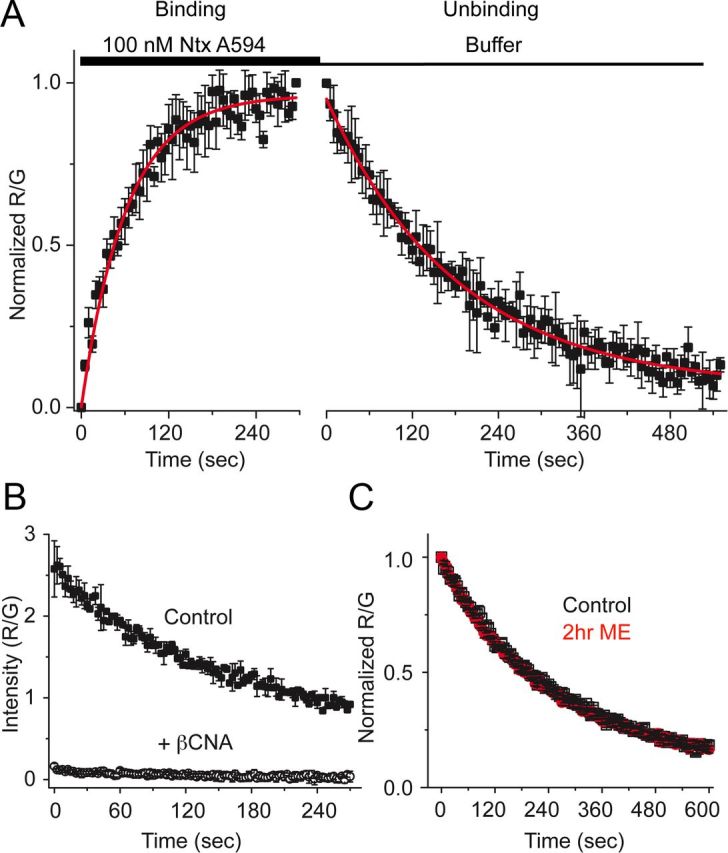
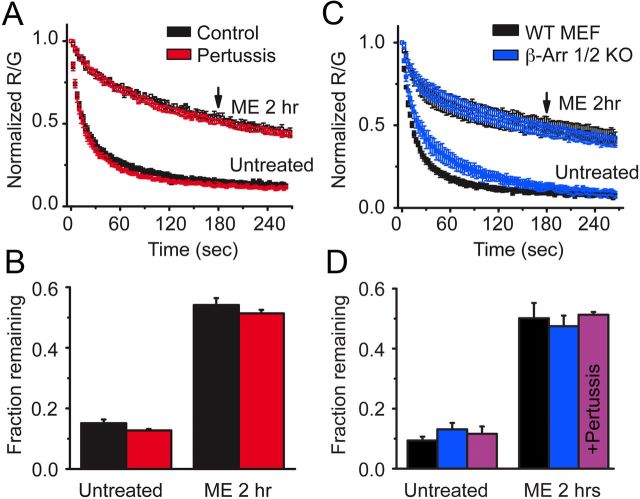
Prolonged exposure to high-efficacy agonists results in desensitization of the μ-opioid receptor (MOR). Desensitized receptors are thought to be unable to couple to G-proteins, preventing downstream signaling; however, the changes to the receptor itself are not well characterized. In the current study, confocal imaging was used to determine whether desensitizing conditions cause a change in agonist-receptor interactions. Using rapid solution exchange, the binding kinetics of fluorescently labeled opioid agonist, dermorphin Alexa594 (derm A594), to MORs was measured in live cells. The affinity of derm A594 binding increased after prolonged treatment of cells with multiple agonists that are known to cause receptor desensitization. In contrast, binding of a fluorescent antagonist, naltrexamine Alexa594, was unaffected by similar agonist pretreatment. The increased affinity of derm A594 for the receptor was long-lived and partially reversed after a 45 min wash. Treatment of the cells with pertussis toxin did not alter the increase in affinity of the derm A594 for MOR. Likewise, the affinity of derm A594 for MORs expressed in mouse embryonic fibroblasts derived from arrestin 1 and 2 knock-out animals increased after treatment of the cells with the desensitization protocol. Thus, opioid receptors were "imprinted" with a memory of prior agonist exposure that was independent of G-protein activation or arrestin binding that altered subsequent agonist-receptor interactions. The increased affinity suggests that acute desensitization results in a long-lasting but reversible conformational change in the receptor.
Figures
References
-
- Ahles A, Rochais F, Frambach T, Bünemann M, Engelhardt S. A polymorphism-specific “memory” mechanism in the β(2)-adrenergic receptor. Sci Signal. 2011;4:ra53. - PubMed
-
- Arttamangkul S, Alvarez-Maubecin V, Thomas G, Williams JT, Grandy DK. Binding and internalization of fluorescent opioid peptide conjugates in living cells. Mol Pharmacol. 2000;58:1570–1580. - PubMed
-
- Benovic JL, Kühn H, Weyand I, Codina J, Caron MG, Lefkowitz RJ. Functional desensitization of the isolated β-adrenergic receptor by the β-adrenergic receptor kinase: potential role of an analog of the retinal protein arrestin (48-kDa protein) Proc Natl Acad Sci U S A. 1987;84:8879–8882. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials