

Cellular differentiation and individuality in the 'minor' multicellular taxa
- PMID: 23448295
- PMCID: PMC4103886
- DOI: 10.1111/brv.12031
Cellular differentiation and individuality in the 'minor' multicellular taxa
Abstract
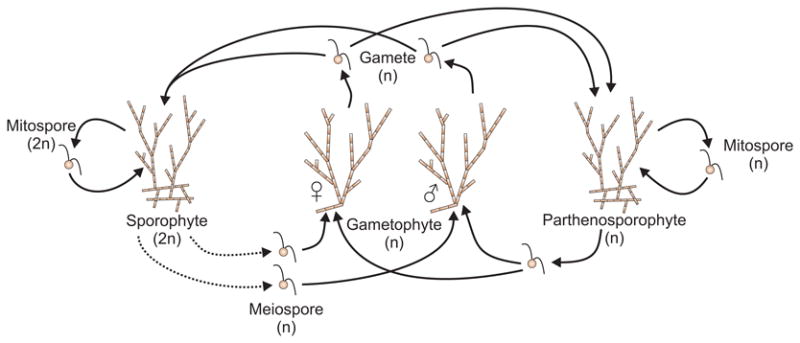
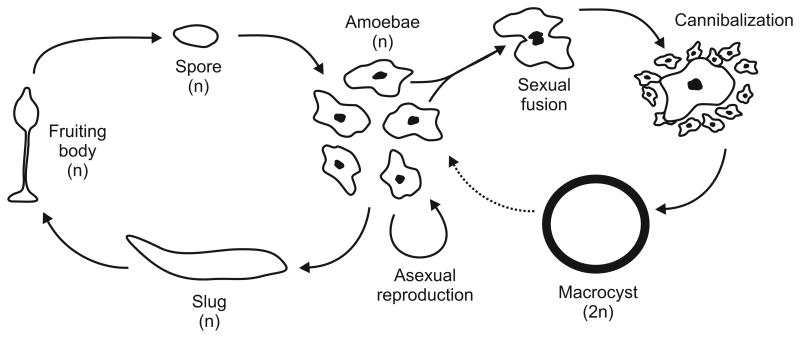
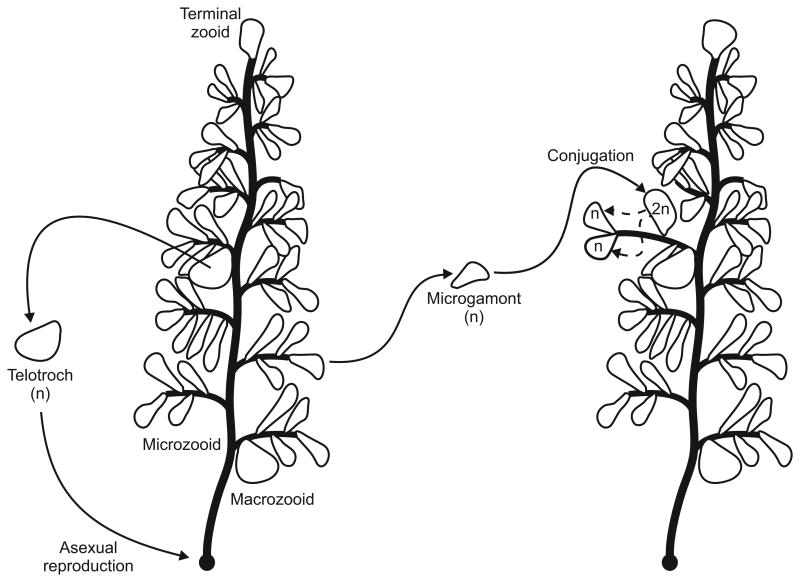
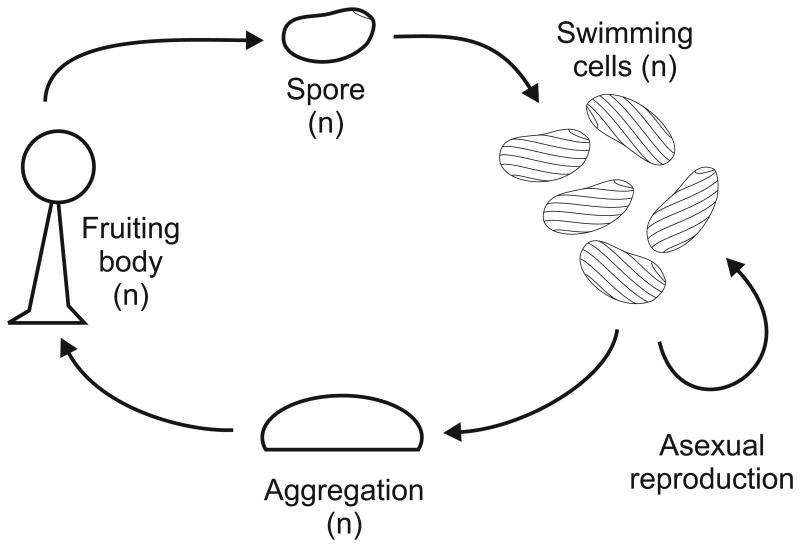
Biology needs a concept of individuality in order to distinguish organisms from parts of organisms and from groups of organisms, to count individuals and compare traits across taxa, and to distinguish growth from reproduction. Most of the proposed criteria for individuality were designed for 'unitary' or 'paradigm' organisms: contiguous, functionally and physiologically integrated, obligately sexually reproducing multicellular organisms with a germ line sequestered early in development. However, the vast majority of the diversity of life on Earth does not conform to all of these criteria. We consider the issue of individuality in the 'minor' multicellular taxa, which collectively span a large portion of the eukaryotic tree of life, reviewing their general features and focusing on a model species for each group. When the criteria designed for unitary organisms are applied to other groups, they often give conflicting answers or no answer at all to the question of whether or not a given unit is an individual. Complex life cycles, intimate bacterial symbioses, aggregative development, and strange genetic features complicate the picture. The great age of some of the groups considered shows that 'intermediate' forms, those with some but not all of the traits traditionally associated with individuality, cannot reasonably be considered ephemeral or assumed transitional. We discuss a handful of recent attempts to reconcile the many proposed criteria for individuality and to provide criteria that can be applied across all the domains of life. Finally, we argue that individuality should be defined without reference to any particular taxon and that understanding the emergence of new kinds of individuals requires recognizing individuality as a matter of degree.
Keywords: cellular differentiation; individuality; life history; major transitions; multicellularity; organisms; symbiosis.
© 2013 The Authors. Biological Reviews © 2013 Cambridge Philosophical Society.
Figures



References
-
- Agutter PS, Tuszynski JA. Analytic theories of allometric scaling. Journal of Experimental Biology. 2011;214:1055–1062. - PubMed
-
- Ariew A, Lewontin RC. The confusions of fitness. British Journal for the Philosopy of Science. 2004;55:347–363.
-
- Ayre DJ, Grosberg RK. Behind anemone lines: factors affecting division of labour in the social cnidarian Anthopleura elegantissima. Animal Behaviour. 2005;70:97–110.
-
- Baldauf SL. The deep roots of eukaryotes. Science. 2003;300:1703–1706. - PubMed
-
- Baldauf SL. An overview of the phylogeny and diversity of eukaryotes. Journal of Systematic Evolution. 2008;46:263–273.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
