

Functional analysis of the murine coronavirus genomic RNA packaging signal
- PMID: 23449786
- PMCID: PMC3624306
- DOI: 10.1128/JVI.00100-13
Functional analysis of the murine coronavirus genomic RNA packaging signal
Abstract
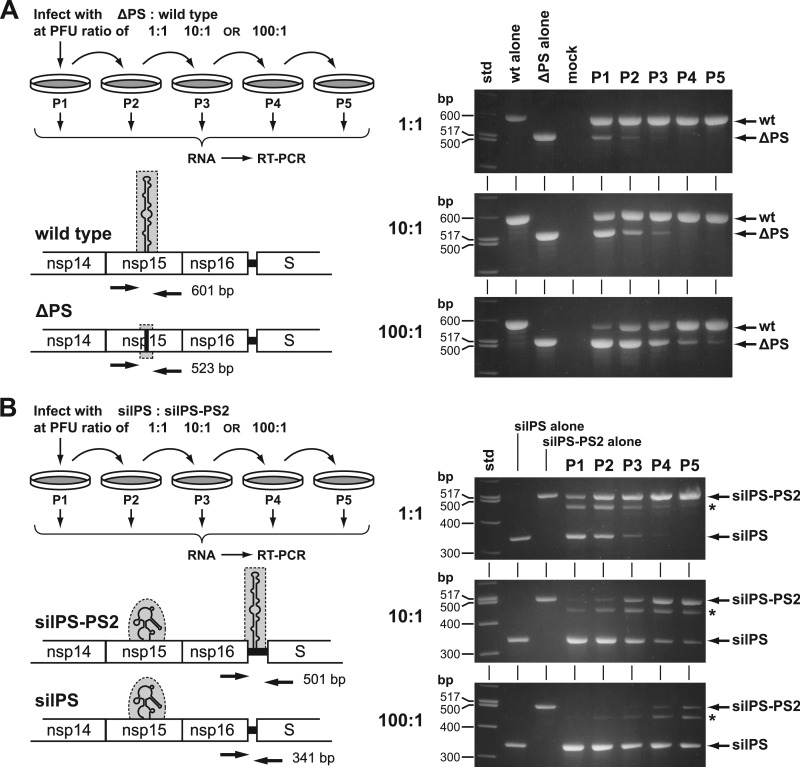
Coronaviruses selectively package genomic RNA into assembled virions, despite the great molar excess of subgenomic RNA species that is present in infected cells. The genomic packaging signal (PS) for the coronavirus mouse hepatitis virus (MHV) was originally identified as an element that conferred packaging capability to defective interfering RNAs. The MHV PS is an RNA structure that maps to the region of the replicase gene encoding the nonstructural protein 15 subunit of the viral replicase-transcriptase complex. To begin to understand the role and mechanism of action of the MHV PS in its native genomic locus, we constructed viral mutants in which this cis-acting element was altered, deleted, or transposed. Our results demonstrated that the PS is pivotal in the selection of viral genomic RNA for incorporation into virions. Mutants in which PS RNA secondary structure was disrupted or entirely ablated packaged large quantities of subgenomic RNAs, in addition to genomic RNA. Moreover, the PS retained its function when displaced to an ectopic site in the genome. Surprisingly, the PS was not essential for MHV viability, nor did its elimination have a severe effect on viral growth. However, the PS was found to provide a distinct selective advantage to MHV. Viruses containing the PS readily outcompeted their otherwise isogenic counterparts lacking the PS.
Figures







References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
