

Unequal recombination and evolution of the mating-type (MAT) loci in the pathogenic fungus Grosmannia clavigera and relatives
- PMID: 23450093
- PMCID: PMC3583454
- DOI: 10.1534/g3.112.004986
Unequal recombination and evolution of the mating-type (MAT) loci in the pathogenic fungus Grosmannia clavigera and relatives
Abstract
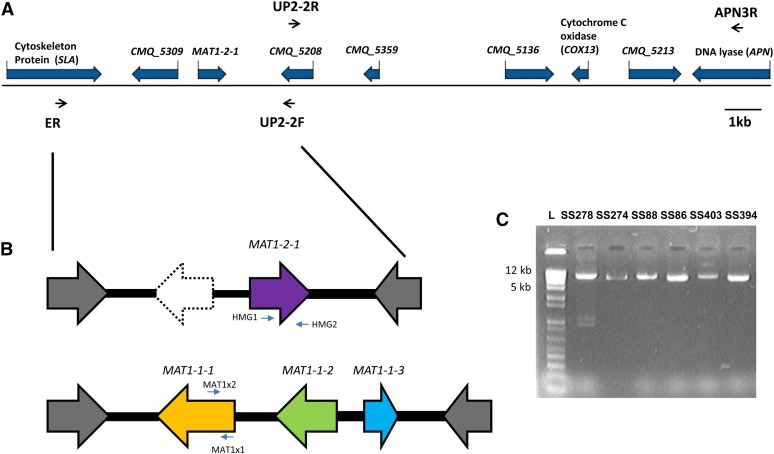
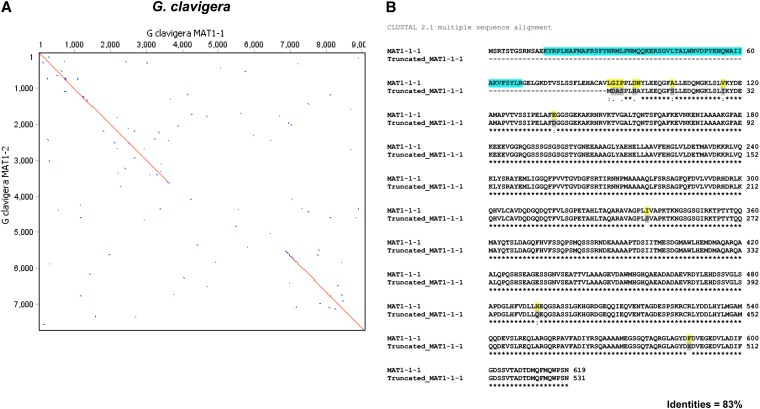
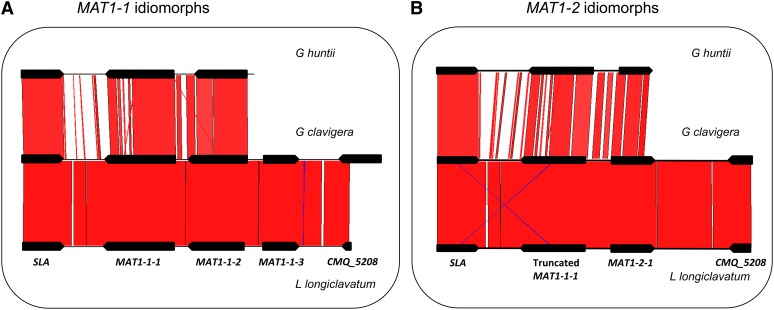
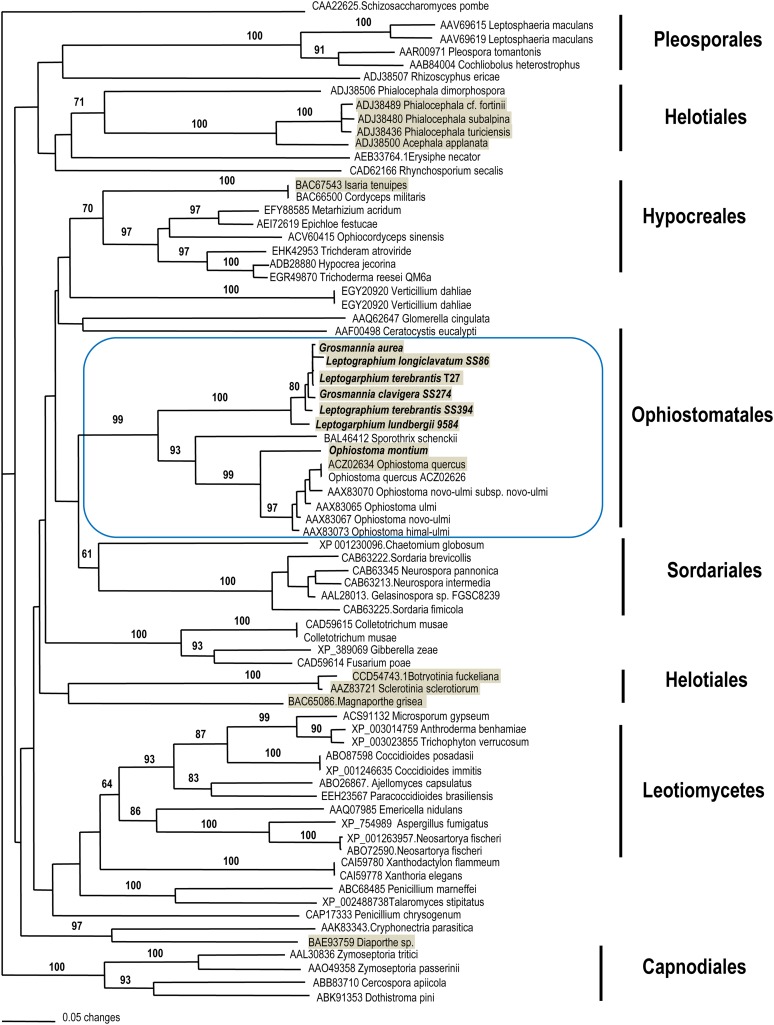
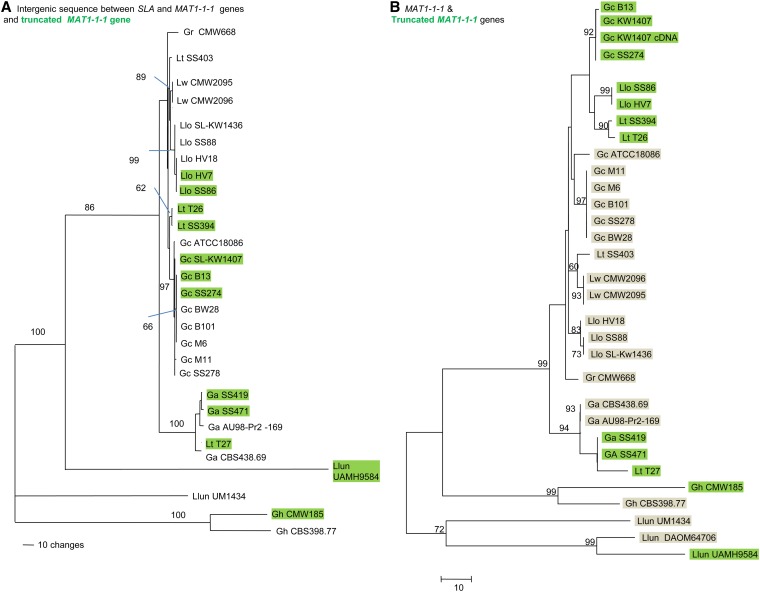
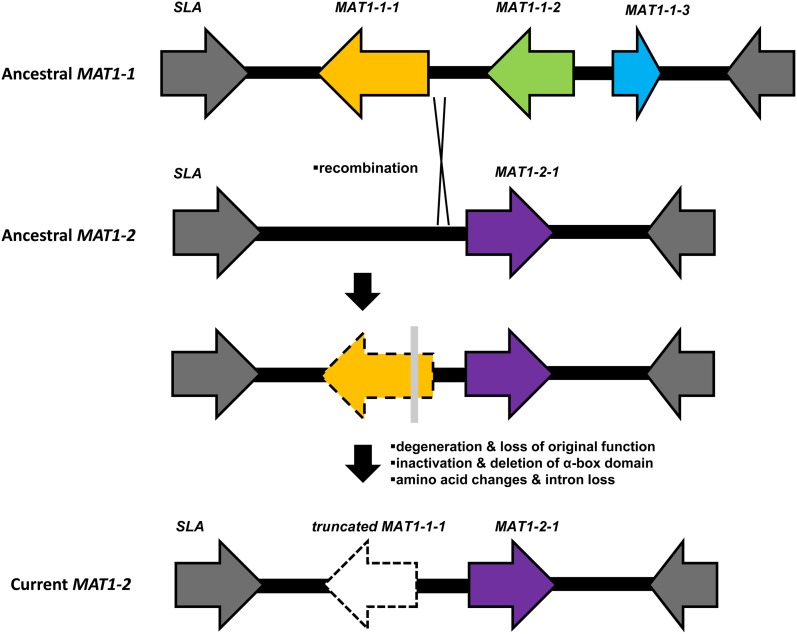
Sexual reproduction in fungi is regulated by the mating-type (MAT) locus where recombination is suppressed. We investigated the evolution of MAT loci in eight fungal species belonging to Grosmannia and Ophiostoma (Sordariomycetes, Ascomycota) that include conifer pathogens and beetle symbionts. The MAT1-2 idiomorph/allele was identified from the assembled and annotated Grosmannia clavigera genome, and the MAT locus is flanked by genes coding for cytoskeleton protein (SLA) and DNA lyase. The synteny of these genes is conserved and consistent with other members in Ascomycota. Using sequences from SLA and flanking regions, we characterized the MAT1-1 idiomorph from other isolates of G. clavigera and performed dotplot analysis between the two idiomorphs. Unexpectedly, the MAT1-2 idiomorph contains a truncated MAT1-1-1 gene upstream of the MAT1-2-1 gene that bears the high-mobility-group domain. The nucleotide and amino acid sequence of the truncated MAT1-1-1 gene is similar to its homologous copy in the MAT1-1 idiomorph in the opposite mating-type isolate, except that positive selection is acting on the truncated gene and the alpha(α)-box that encodes the transcription factor has been deleted. The MAT idiomorphs sharing identical gene organization were present in seven additional species in the Ophiostomatales, suggesting that the presence of truncated MAT1-1-1 gene is a general pattern in this order. We propose that an ancient unequal recombination event resulted in the ancestral MAT1-1-1 gene integrated into the MAT1-2 idiomorph and surviving as the truncated MAT1-1-1 genes. The α-box domain of MAT1-1-1 gene, located at the same MAT locus adjacent to the MAT1-2-1 gene, could have been removed by deletion after recombination due to mating signal interference. Our data confirmed a 1:1 MAT/sex ratio in two pathogen populations, and showed that all members of the Ophiostomatales studied here including those that were previously deemed asexual have the potential to reproduce sexually. This ability can potentially increase genetic variability and can enhance fitness in new, ecological niches.
Keywords: heterothallism; homothallism; mating system evolution; outcrossing; selfing.
Figures

References
-
- Alamouti S. M., Wang V., DiGuistini S., Six D., Bohlmann J., et al. , 2011. Gene genealogies reveal cryptic species and host preferences for the pine fungal pathogen Grosmannia clavigera. Mol. Ecol. 12: 2581–2602 - PubMed
-
- Bennett R. S., Yun S., Lee T. Y., Turgeon B. G., Arseniuk E., et al. , 2003. Identity and conservation of mating type genes in geographically diverse isolates of Phaeosphaeria nodorum. Fungal Genet. Biol. 40: 25–37 - PubMed
-
- Billiard S., López-Villavicencio M., Devier B., Hood M. E., Fairhead C., et al. , 2011. Having sex, yes, but with whom? Inferences from fungi on the evolution of anisogamy and mating types. Biol. Rev. Camb. Philos. Soc. 86: 421–442 - PubMed
-
- Billiard S., Lopez-Villavicencio M., Hood M. E., Giraud T., 2012. Sex, outcrossing and mating types: unsolved questions in fungi and beyond. J. Evol. Biol. 25: 1020–1038 - PubMed
Publication types
MeSH terms
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources