

Calcineurin governs thermotolerance and virulence of Cryptococcus gattii
- PMID: 23450261
- PMCID: PMC3583459
- DOI: 10.1534/g3.112.004242
Calcineurin governs thermotolerance and virulence of Cryptococcus gattii
Abstract
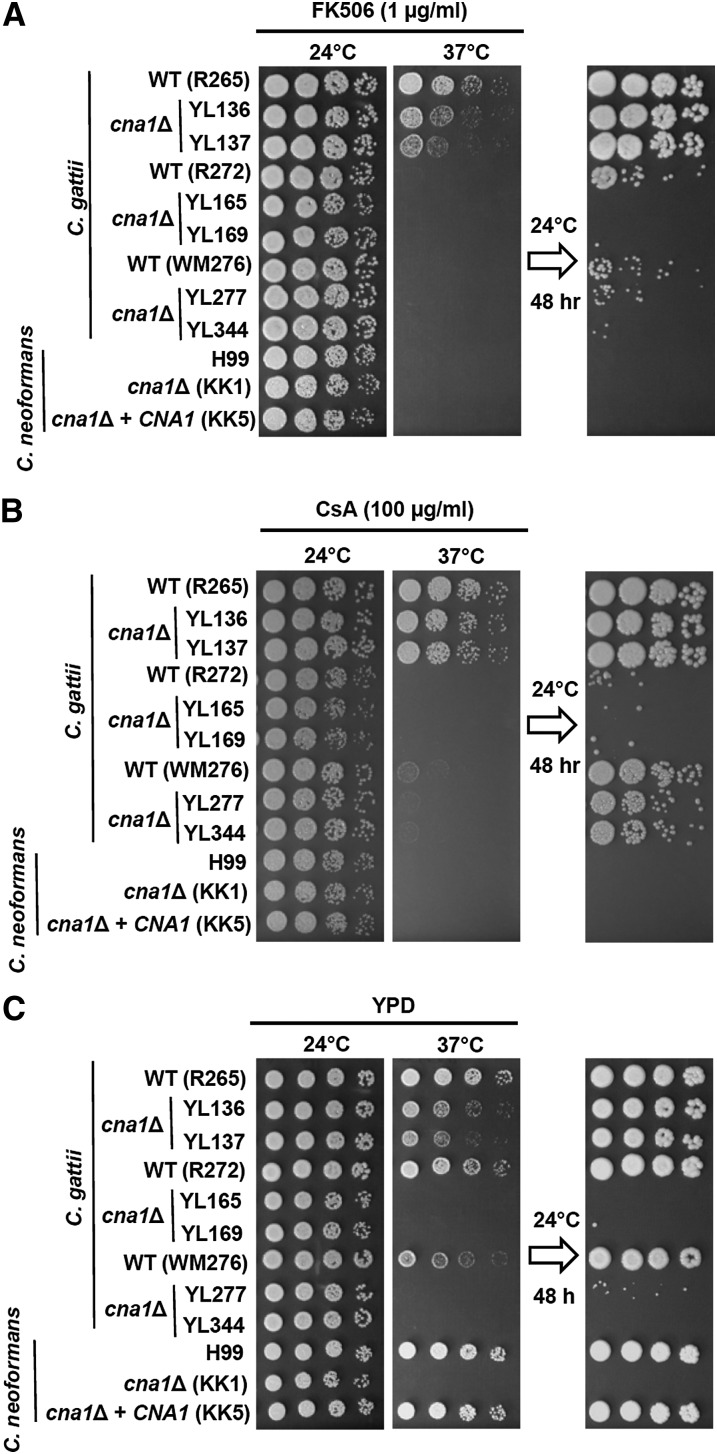
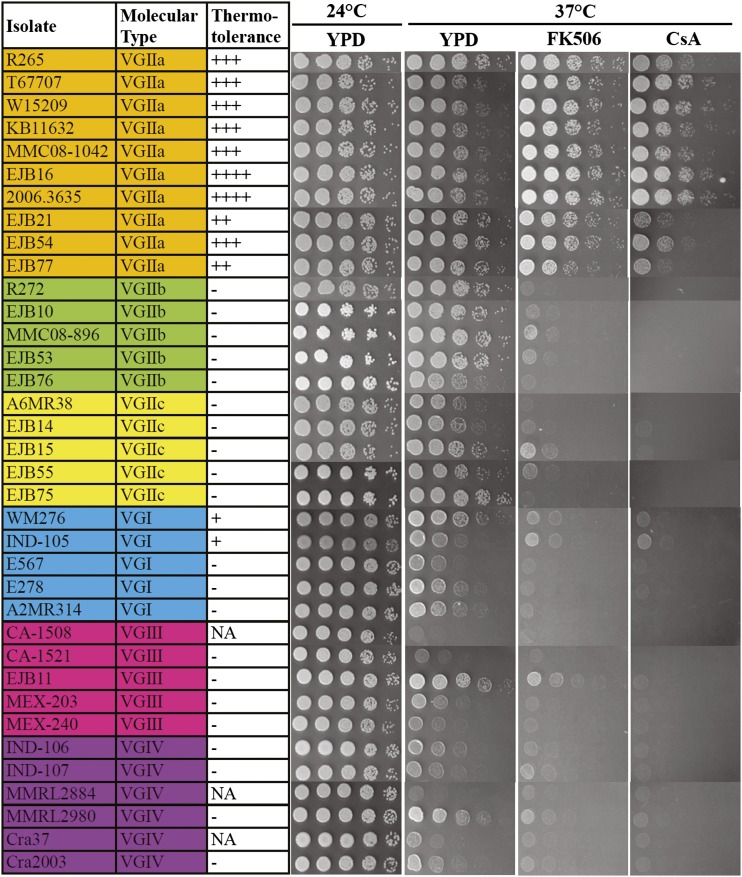
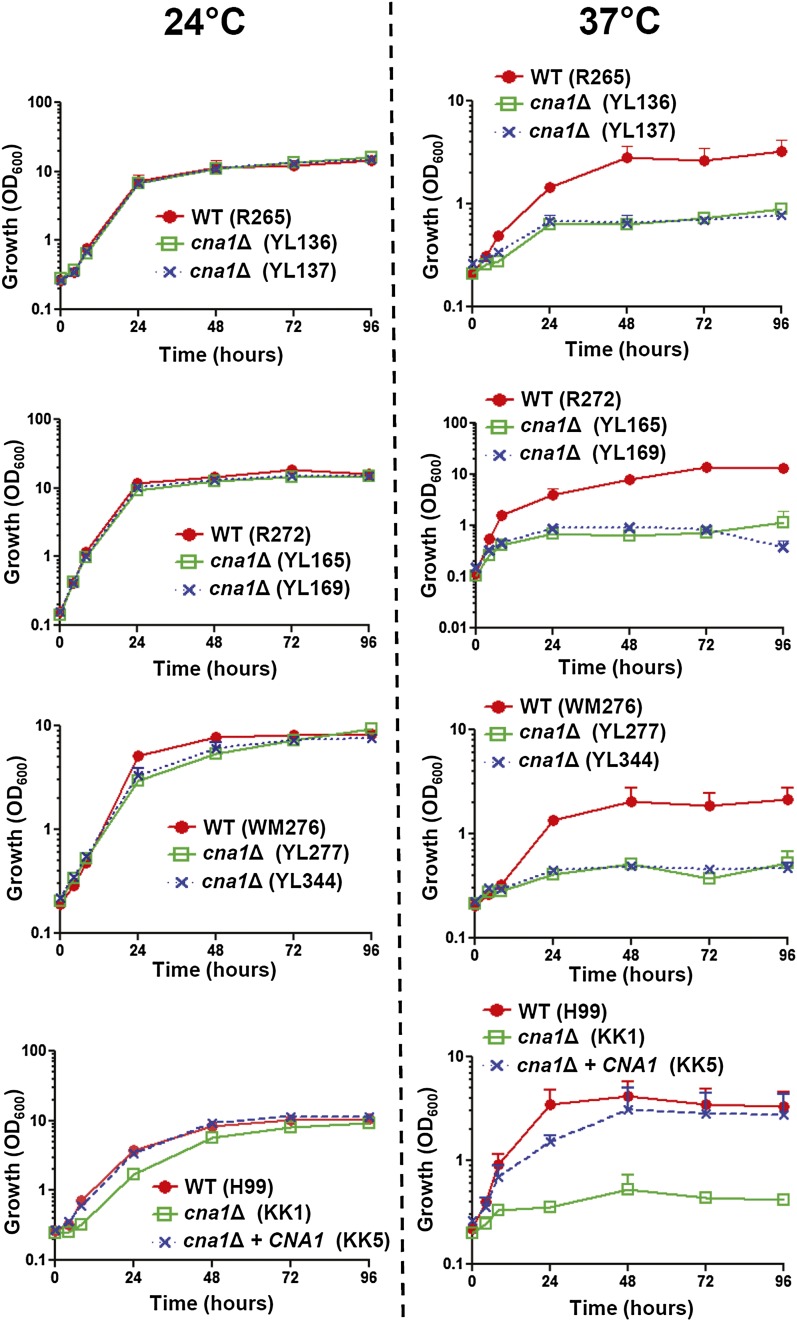
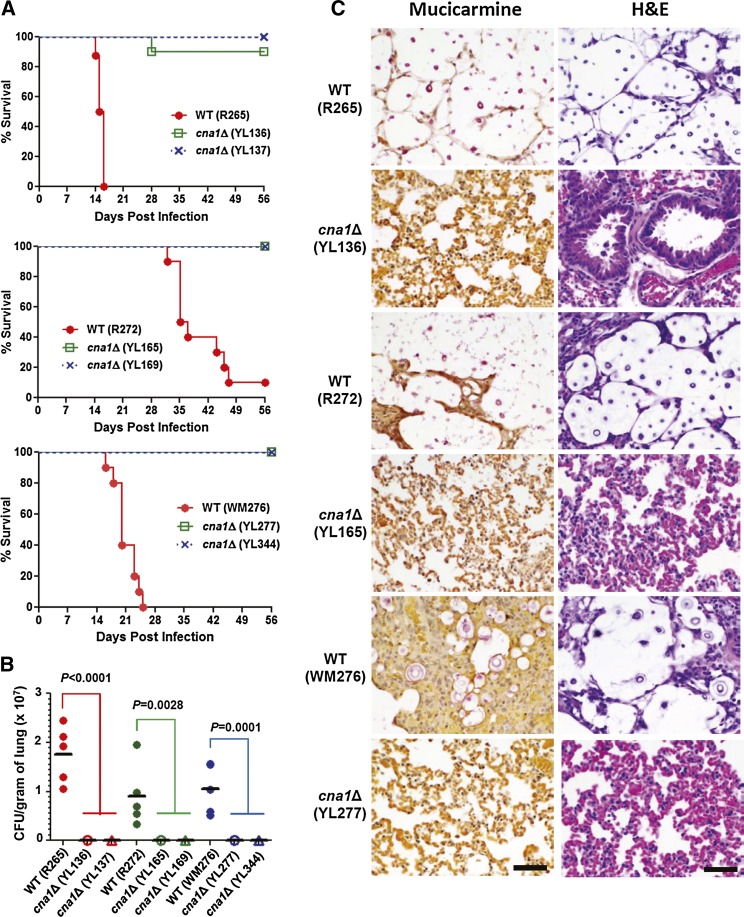
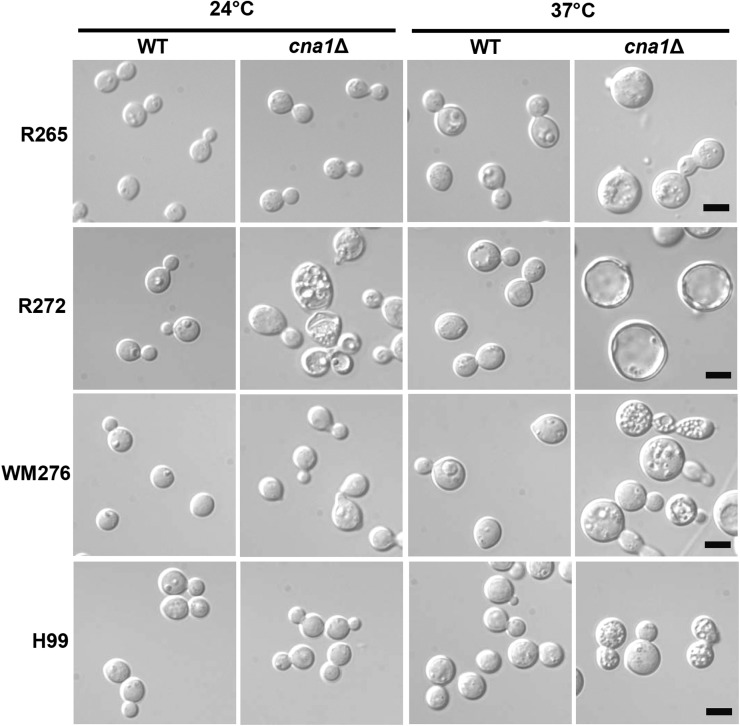
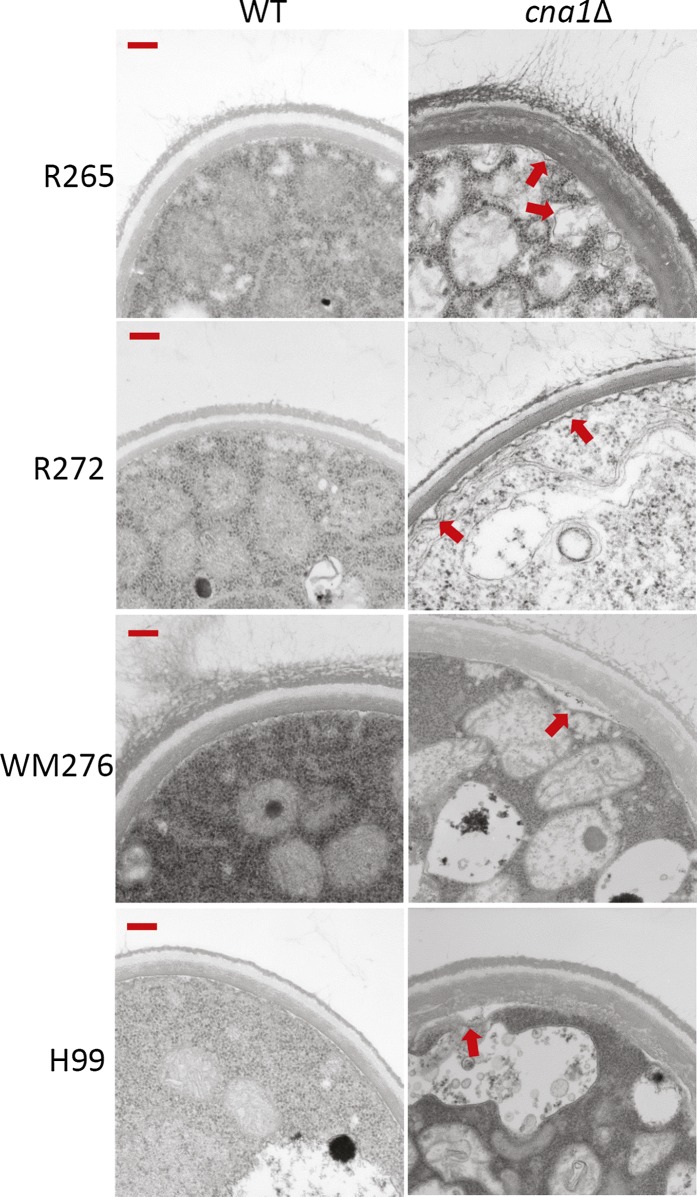
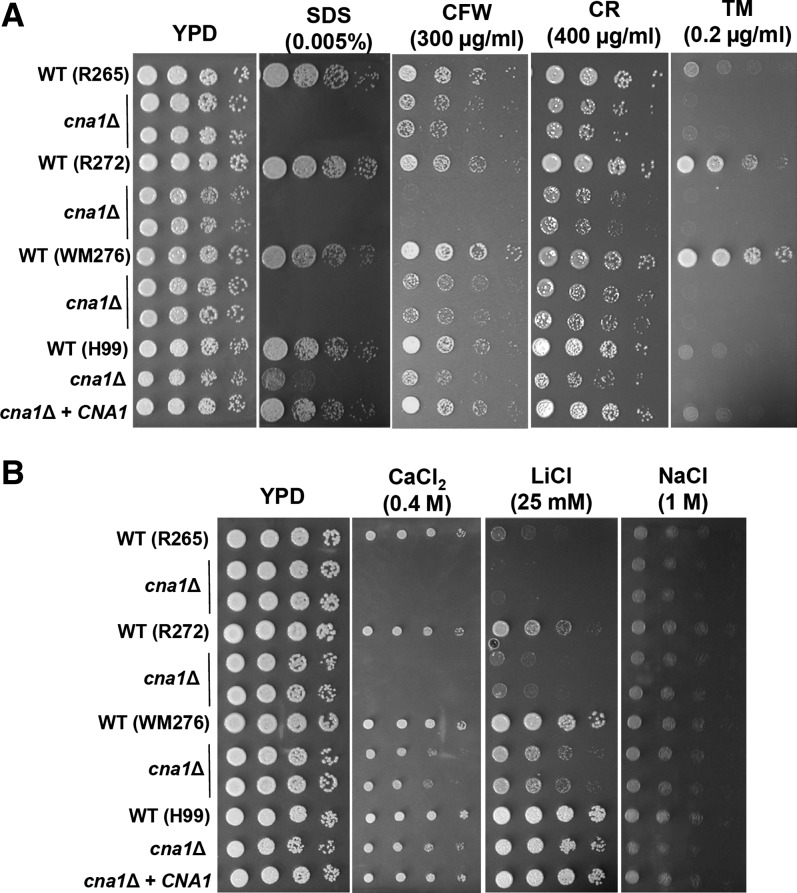
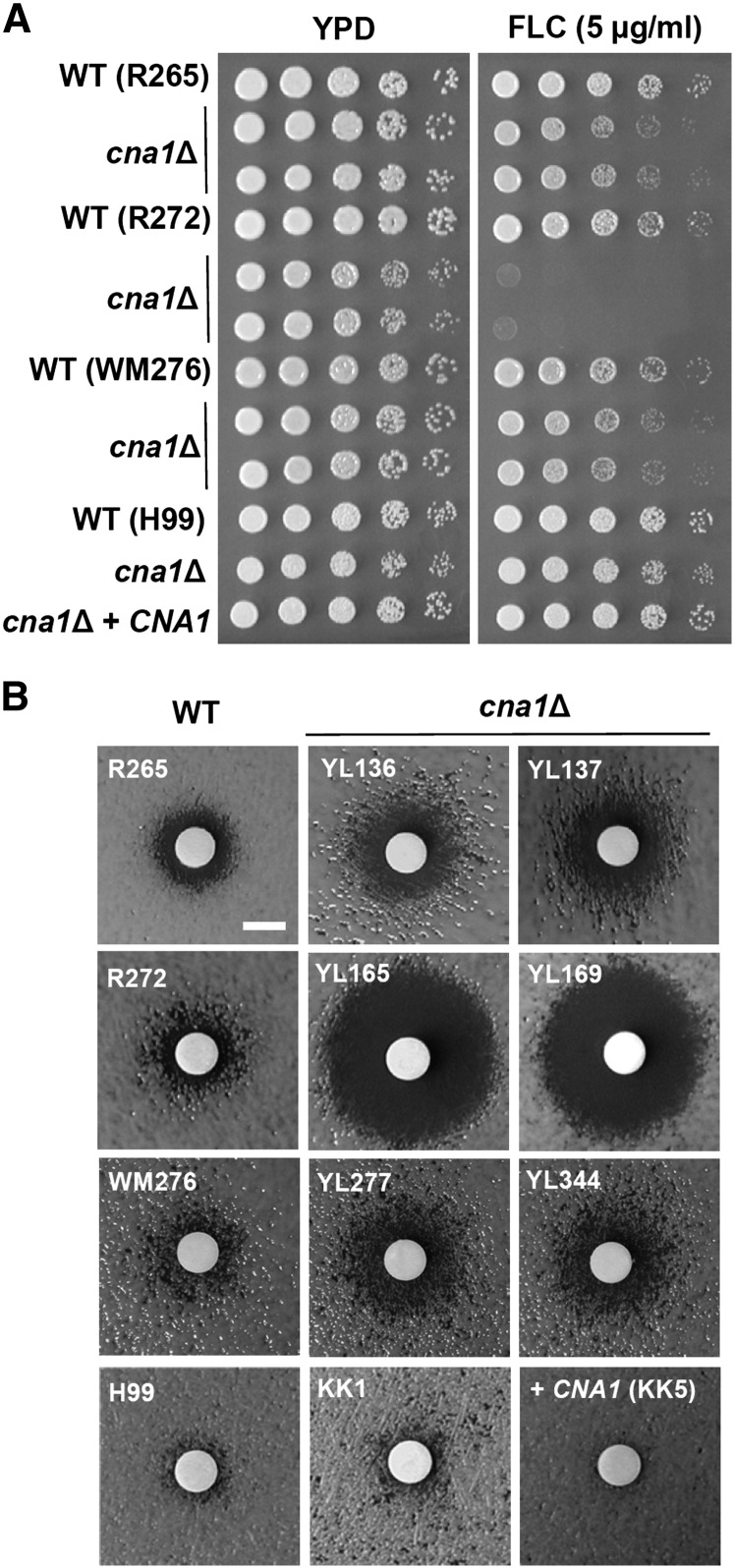
The pathogenic yeast Cryptococcus gattii, which is causing an outbreak in the Pacific Northwest region of North America, causes life-threatening pulmonary infections and meningoencephalitis in healthy individuals, unlike Cryptococcus neoformans, which commonly infects immunocompromised patients. In addition to a greater predilection for C. gattii to infect healthy hosts, the C. gattii genome sequence project revealed extensive chromosomal rearrangements compared with C. neoformans, showing genomic differences between the two Cryptococcus species. We investigated the roles of C. gattii calcineurin in three molecular types: VGIIa (R265), VGIIb (R272), and VGI (WM276). We found that calcineurin exhibits a differential requirement for growth on solid medium at 37°, as calcineurin mutants generated from R265 were more thermotolerant than mutants from R272 and WM276. We demonstrated that tolerance to calcineurin inhibitors (FK506, CsA) at 37° is linked with the VGIIa molecular type. The calcineurin mutants from the R272 background showed the most extensive growth and morphological defects (multivesicle and larger ring-like cells), as well as increased fluconazole susceptibility. Our cellular architecture examination showed that C. gattii and C. neoformans calcineurin mutants exhibit plasma membrane disruptions. Calcineurin in the C. gattii VGII molecular type plays a greater role in controlling cation homeostasis compared with that in C. gattii VGI and C. neoformans H99. Importantly, we demonstrate that C. gattii calcineurin is essential for virulence in a murine inhalation model, supporting C. gattii calcineurin as an attractive antifungal drug target.
Keywords: FK506; capsule; cation homeostasis; cyclosporin A; fluconazole tolerance; melanin; pathogenicity; temperature sensitivity.
Figures
Similar articles
-
Cryptococcus gattii infections.Clin Microbiol Rev. 2014 Oct;27(4):980-1024. doi: 10.1128/CMR.00126-13. Clin Microbiol Rev. 2014. PMID: 25278580 Free PMC article. Review.
-
Cryptococcus neoformans-Cryptococcus gattii species complex: an international study of wild-type susceptibility endpoint distributions and epidemiological cutoff values for fluconazole, itraconazole, posaconazole, and voriconazole.Antimicrob Agents Chemother. 2012 Nov;56(11):5898-906. doi: 10.1128/AAC.01115-12. Epub 2012 Sep 4. Antimicrob Agents Chemother. 2012. PMID: 22948877 Free PMC article.
-
Azole resistance in Cryptococcus gattii from the Pacific Northwest: Investigation of the role of ERG11.Antimicrob Agents Chemother. 2013 Nov;57(11):5478-85. doi: 10.1128/AAC.02287-12. Epub 2013 Aug 26. Antimicrob Agents Chemother. 2013. PMID: 23979758 Free PMC article.
-
Molecular typing, in vitro susceptibility and virulence of Cryptococcus neoformans/Cryptococcus gattii species complex clinical isolates from south-eastern Brazil.Mycoses. 2020 Dec;63(12):1341-1351. doi: 10.1111/myc.13174. Epub 2020 Sep 30. Mycoses. 2020. PMID: 32869413
-
Cryptococcus gattii comparative genomics and transcriptomics: a NIH/NIAID White Paper.Mycopathologia. 2012 Jun;173(5-6):367-73. doi: 10.1007/s11046-011-9512-9. Epub 2011 Dec 17. Mycopathologia. 2012. PMID: 22179781 Review.
Cited by
-
Fungi that Infect Humans.Microbiol Spectr. 2017 Jun;5(3):10.1128/microbiolspec.funk-0014-2016. doi: 10.1128/microbiolspec.FUNK-0014-2016. Microbiol Spectr. 2017. PMID: 28597822 Free PMC article. Review.
-
Cryptococcus gattii infections.Clin Microbiol Rev. 2014 Oct;27(4):980-1024. doi: 10.1128/CMR.00126-13. Clin Microbiol Rev. 2014. PMID: 25278580 Free PMC article. Review.
-
Antifungal Therapy: New Advances in the Understanding and Treatment of Mycosis.Front Microbiol. 2017 Jan 23;8:36. doi: 10.3389/fmicb.2017.00036. eCollection 2017. Front Microbiol. 2017. PMID: 28167935 Free PMC article. Review.
-
The monothiol glutaredoxin Grx4 influences thermotolerance, cell wall integrity, and Mpk1 signaling in Cryptococcus neoformans.G3 (Bethesda). 2021 Oct 19;11(11):jkab322. doi: 10.1093/g3journal/jkab322. G3 (Bethesda). 2021. PMID: 34542604 Free PMC article.
-
A Novel Resistance Pathway for Calcineurin Inhibitors in the Human-Pathogenic Mucorales Mucor circinelloides.mBio. 2020 Jan 28;11(1):e02949-19. doi: 10.1128/mBio.02949-19. mBio. 2020. PMID: 31992620 Free PMC article.
References
-
- Bovers M., Hagen F., Kuramae E. E., Boekhout T., 2008. Six monophyletic lineages identified within Cryptococcus neoformans and Cryptococcus gattii by multi-locus sequence typing. Fungal Genet. Biol. 45: 400–421 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases