

How cholesterol interacts with membrane proteins: an exploration of cholesterol-binding sites including CRAC, CARC, and tilted domains
- PMID: 23450735
- PMCID: PMC3584320
- DOI: 10.3389/fphys.2013.00031
How cholesterol interacts with membrane proteins: an exploration of cholesterol-binding sites including CRAC, CARC, and tilted domains
Abstract
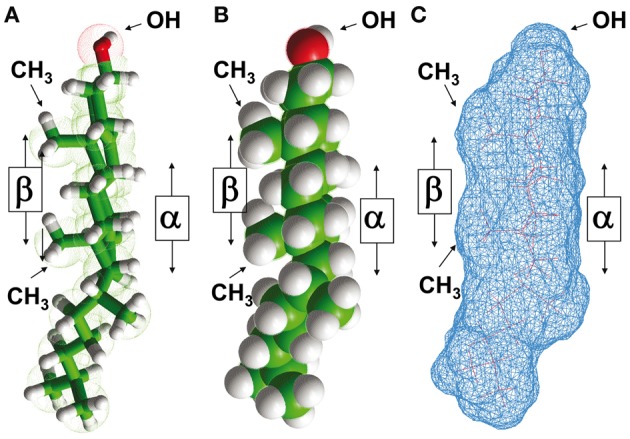
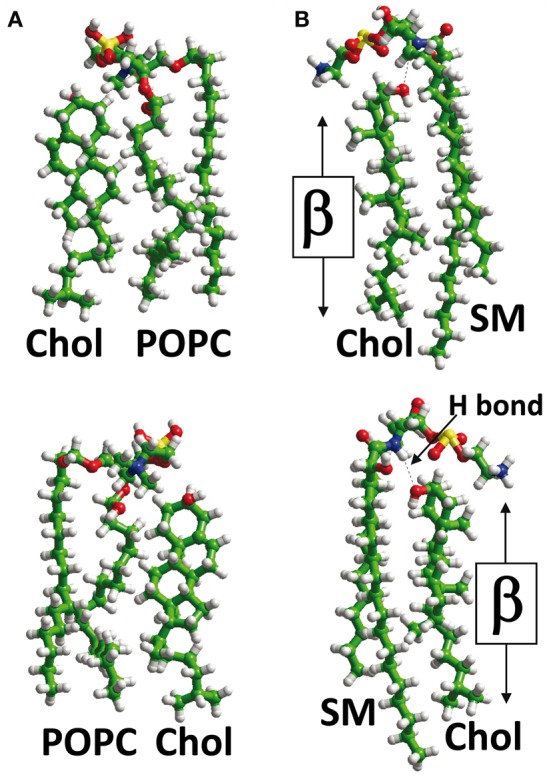
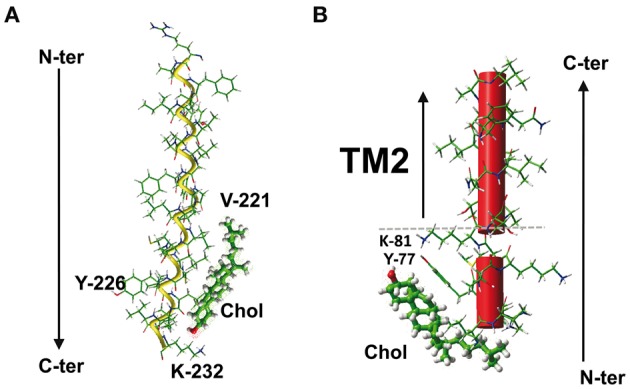
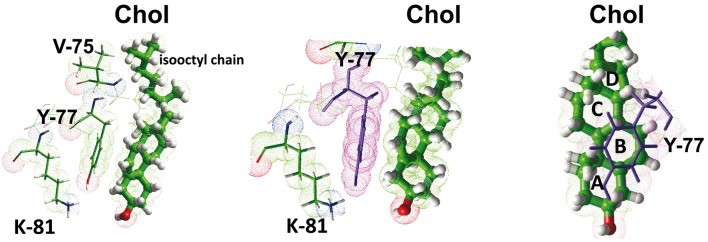
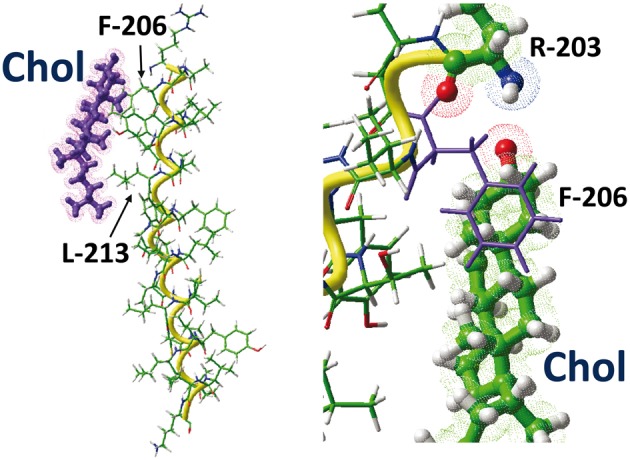
The plasma membrane of eukaryotic cells contains several types of lipids displaying high biochemical variability in both their apolar moiety (e.g., the acyl chain of glycerolipids) and their polar head (e.g., the sugar structure of glycosphingolipids). Among these lipids, cholesterol is unique because its biochemical variability is almost exclusively restricted to the oxidation of its polar -OH group. Although generally considered the most rigid membrane lipid, cholesterol can adopt a broad range of conformations due to the flexibility of its isooctyl chain linked to the polycyclic sterane backbone. Moreover, cholesterol is an asymmetric molecule displaying a planar α face and a rough β face. Overall, these structural features open up a number of possible interactions between cholesterol and membrane lipids and proteins, consistent with the prominent regulatory functions that this unique lipid exerts on membrane components. The aim of this review is to describe how cholesterol interacts with membrane lipids and proteins at the molecular/atomic scale, with special emphasis on transmembrane domains of proteins containing either the consensus cholesterol-binding motifs CRAC and CARC or a tilted peptide. Despite their broad structural diversity, all these domains bind cholesterol through common molecular mechanisms, leading to the identification of a subset of amino acid residues that are overrepresented in both linear and three-dimensional membrane cholesterol-binding sites.
Keywords: Alzheimer; CH-Pi; alpha-synuclein; cholesterol; lipid raft; lipid-protein interaction; neurotransmitter; receptor structure.
Figures



References
-
- Ballesteros J., Weinstein H. (1995). Integrated methods for the construction of three-dimensional models and computational probing of structure-function relations in G protein-coupled receptors. Methods Neurosci. 25, 366–428
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous