

Molecular traffic jams on DNA
- PMID: 23451891
- PMCID: PMC3651777
- DOI: 10.1146/annurev-biophys-083012-130304
Molecular traffic jams on DNA
Abstract
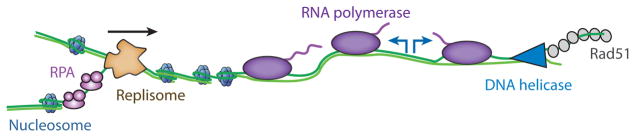
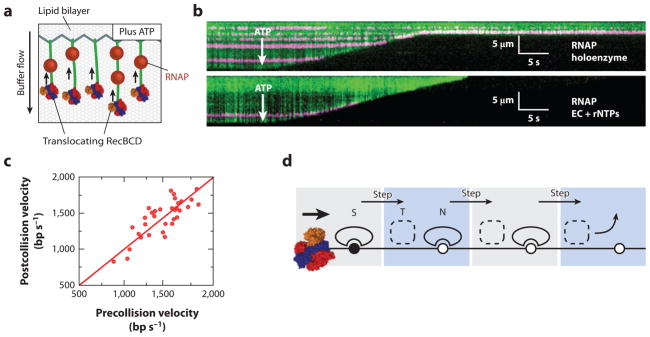
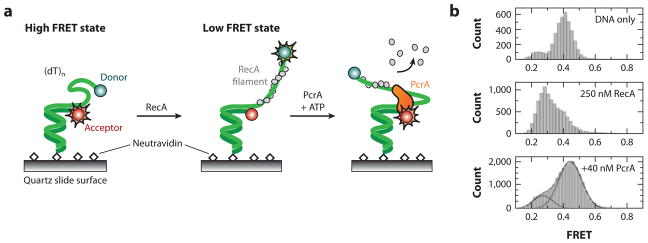
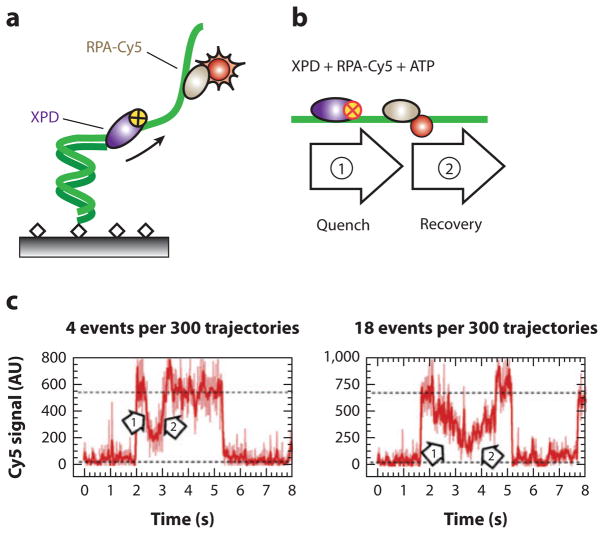
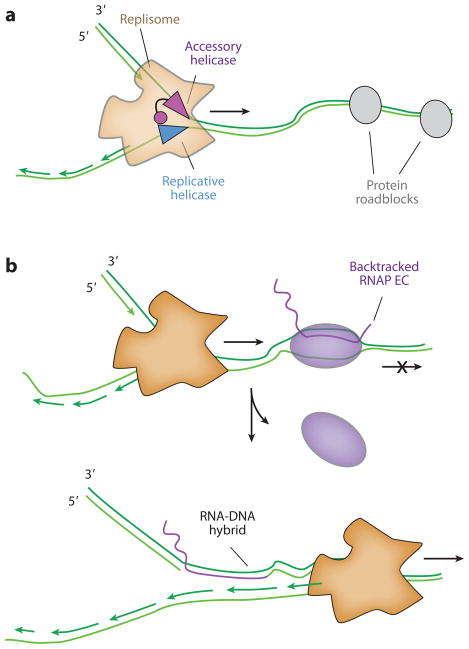
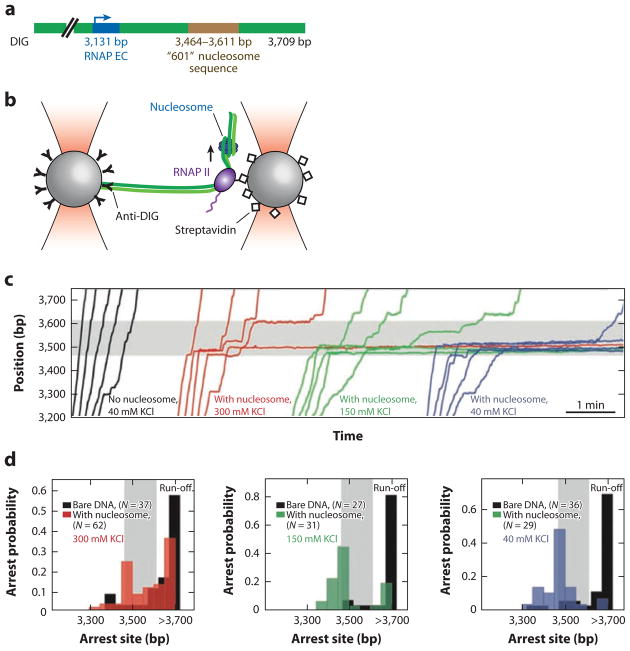
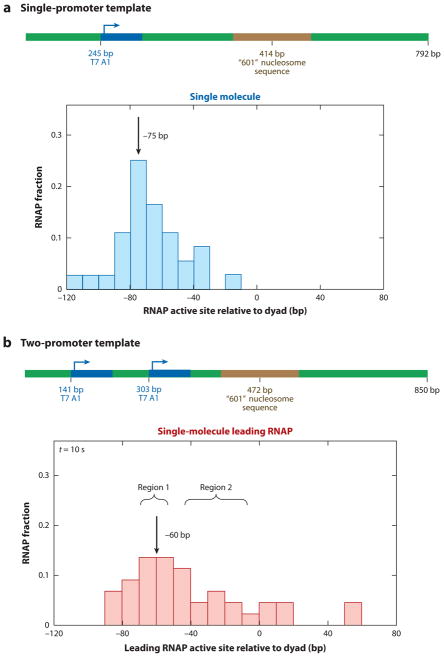
All aspects of DNA metabolism-including transcription, replication, and repair-involve motor enzymes that move along genomic DNA. These processes must all take place on chromosomes that are occupied by a large number of other proteins. However, very little is known regarding how nucleic acid motor proteins move along the crowded DNA substrates that are likely to exist in physiological settings. This review summarizes recent progress in understanding how DNA-binding motor proteins respond to the presence of other proteins that lie in their paths. We highlight recent single-molecule biophysical experiments aimed at addressing this question, with an emphasis placed on analyzing the single-molecule, ensemble biochemical, and in vivo data from a mechanistic perspective.
Figures
References
-
- Aguilera A, García-Muse T. R Loops: from transcription byproducts to threats to genome stability. Mol Cell. 2012;46:115–24. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
