

Considering protonation as a posttranslational modification regulating protein structure and function
- PMID: 23451893
- PMCID: PMC4041481
- DOI: 10.1146/annurev-biophys-050511-102349
Considering protonation as a posttranslational modification regulating protein structure and function
Abstract
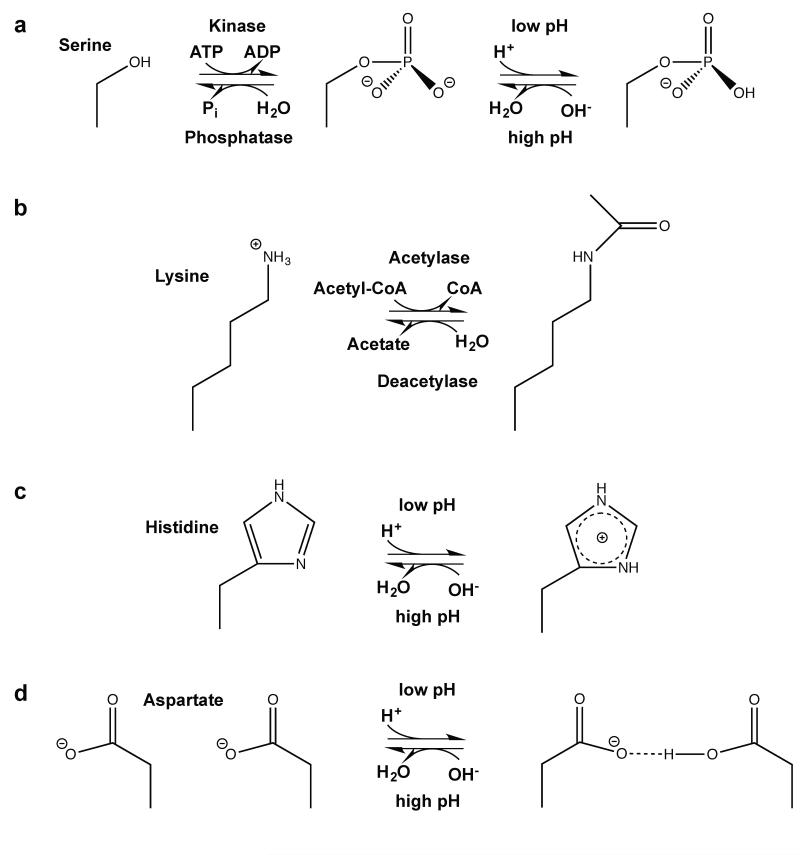
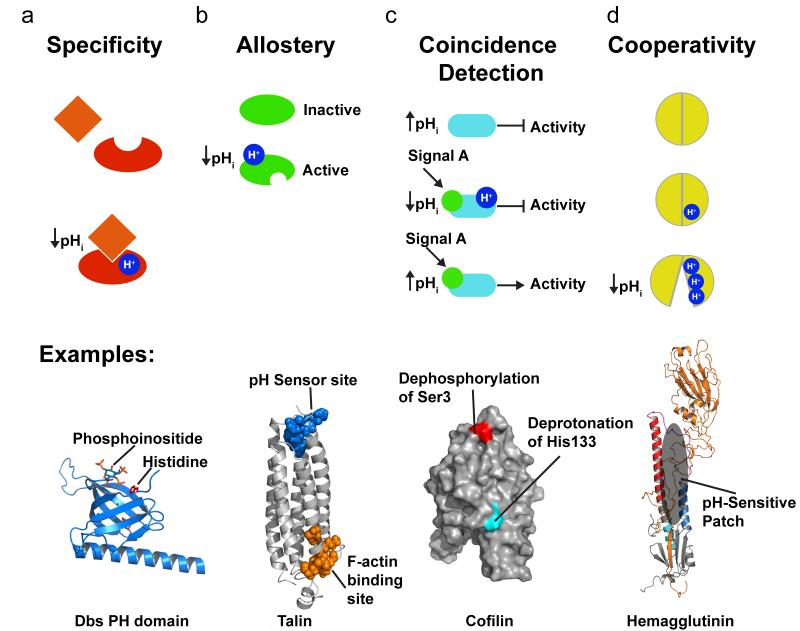
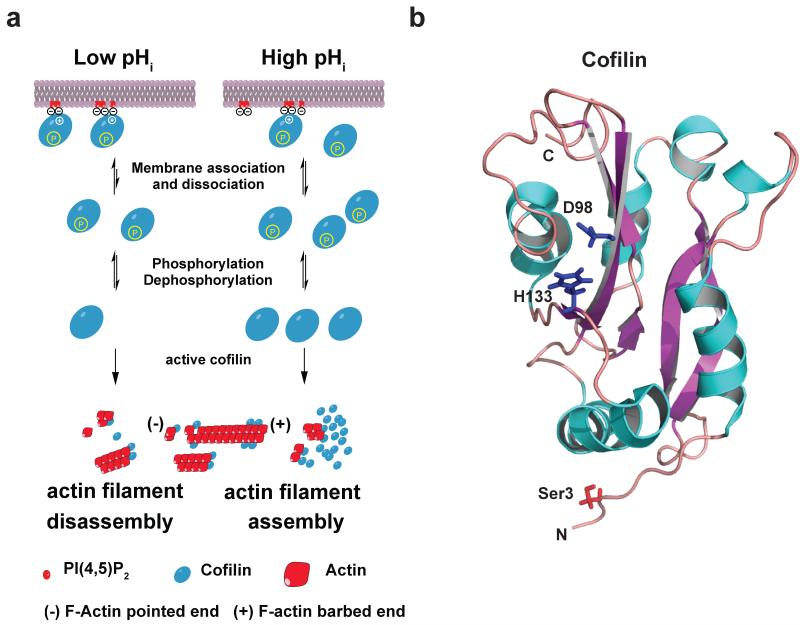
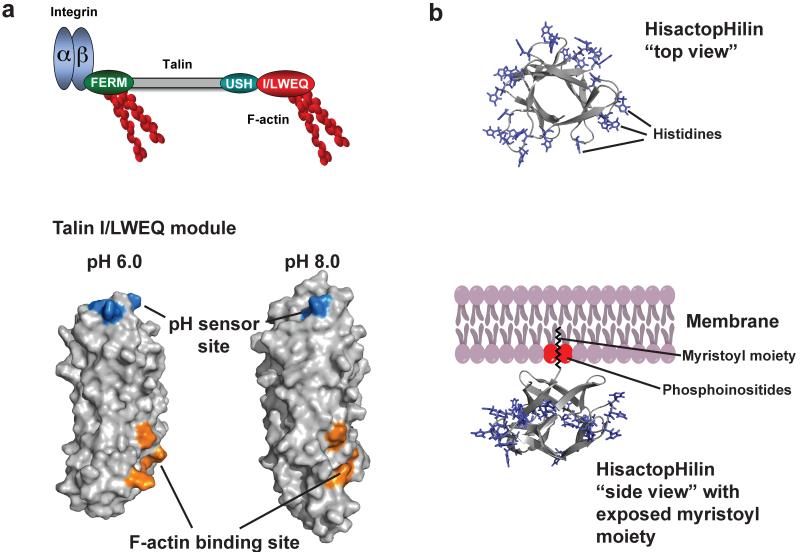
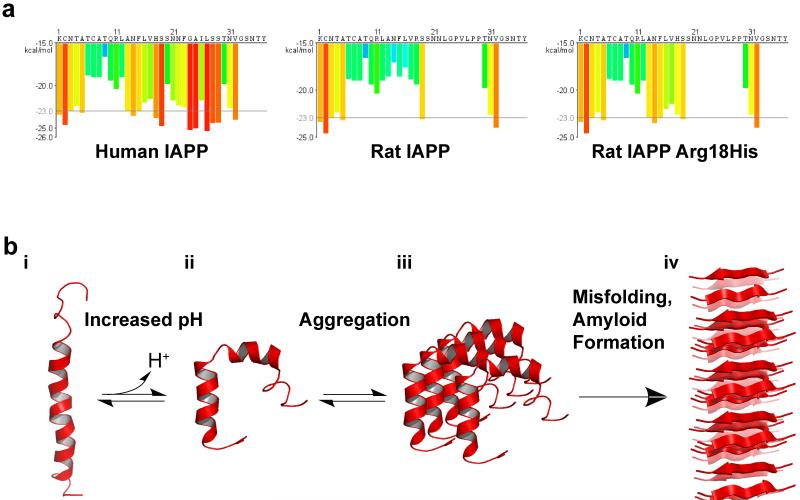
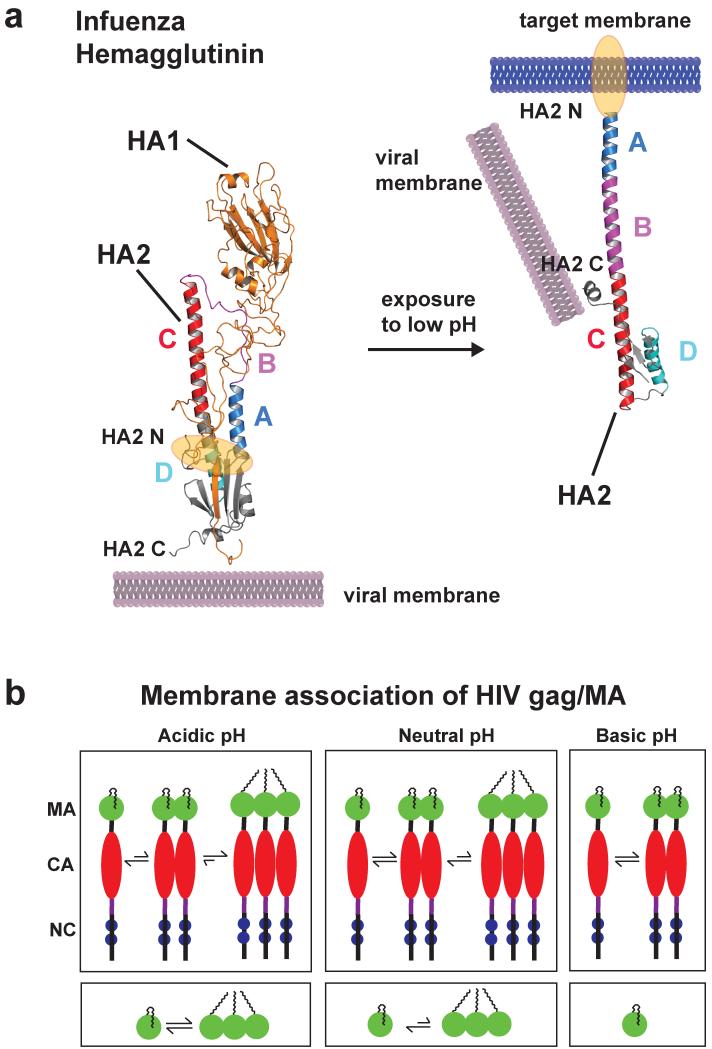
Posttranslational modification is an evolutionarily conserved mechanism for regulating protein activity, binding affinity, and stability. Compared with established posttranslational modifications such as phosphorylation or ubiquitination, posttranslational modification by protons within physiological pH ranges is a less recognized mechanism for regulating protein function. By changing the charge of amino acid side chains, posttranslational modification by protons can drive dynamic changes in protein conformation and function. Addition and removal of a proton is rapid and reversible and, in contrast to most other posttranslational modifications, does not require an enzyme. Signaling specificity is achieved by only a minority of sites in proteins titrating within the physiological pH range. Here, we examine the structural mechanisms and functional consequences of proton posttranslational modification of pH-sensing proteins regulating different cellular processes.
Figures
References
-
- Andrianantoandro E, Pollard TD. Mechanism of actin filament turnover by severing and nucleation at different concentrations of ADF/cofilin. Molecular cell. 2006;24:13–23. - PubMed
-
- Baddam S, Bowler BE. Conformationally gated electron transfer in iso-1-cytochrome c: engineering the rate of a conformational switch. J Am Chem Soc. 2005;127:9702–3. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
