

Characterization of a pre-export enzyme-chaperone complex on the twin-arginine transport pathway
- PMID: 23452237
- PMCID: PMC3635657
- DOI: 10.1042/BJ20121832
Characterization of a pre-export enzyme-chaperone complex on the twin-arginine transport pathway
Abstract
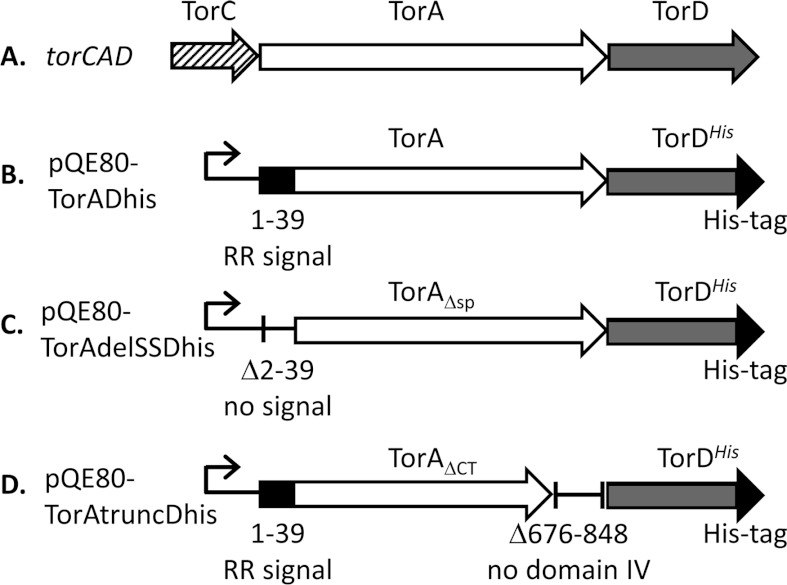
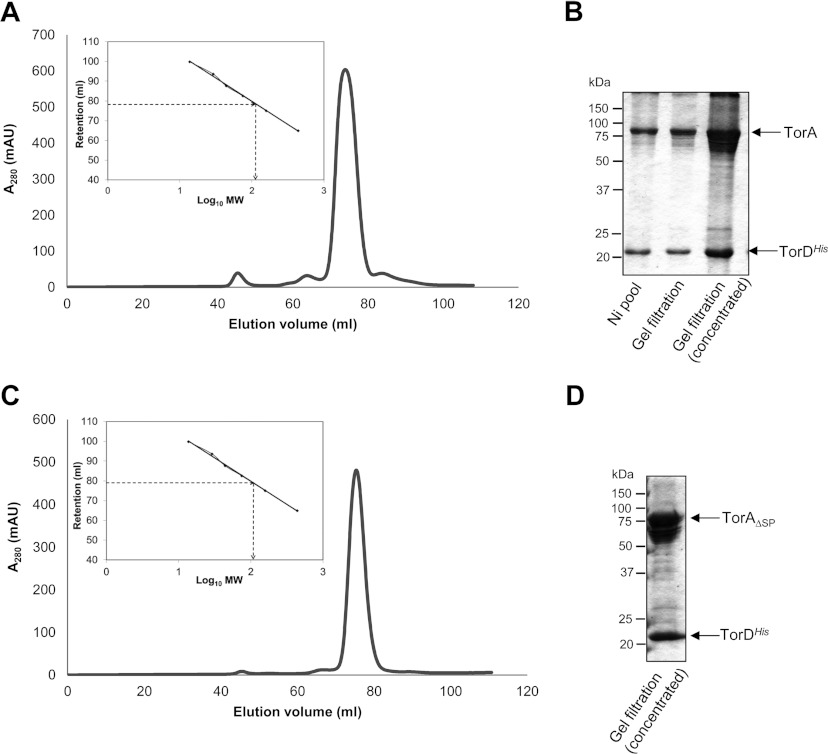
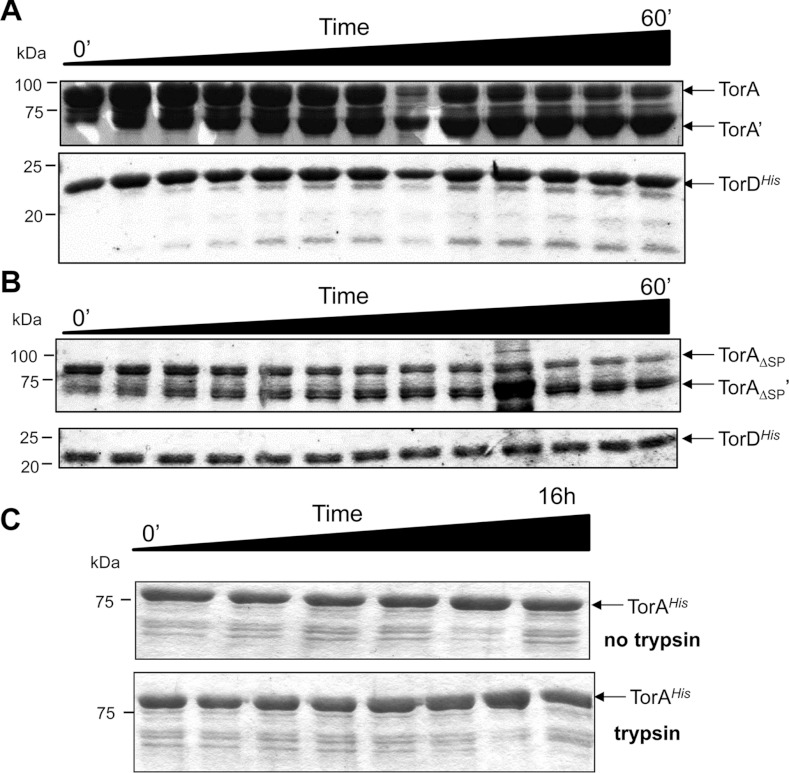
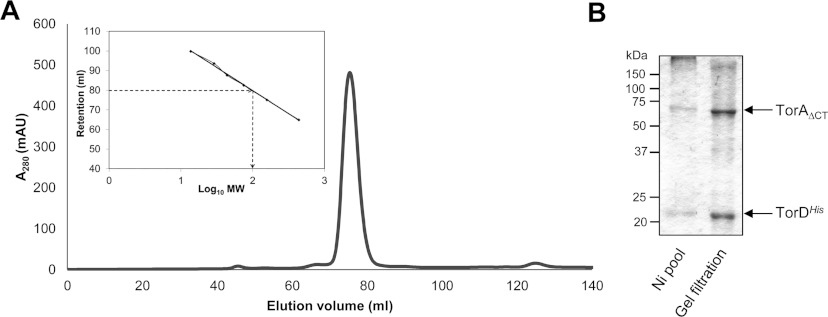
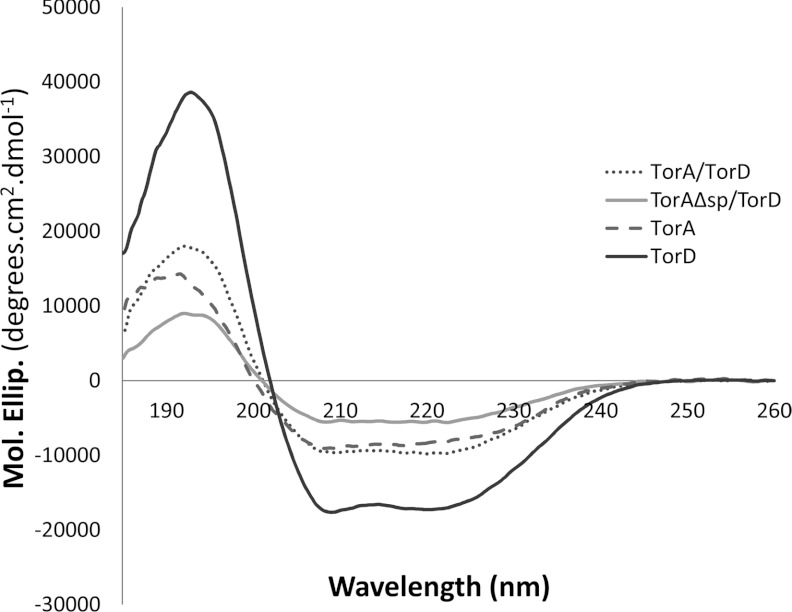
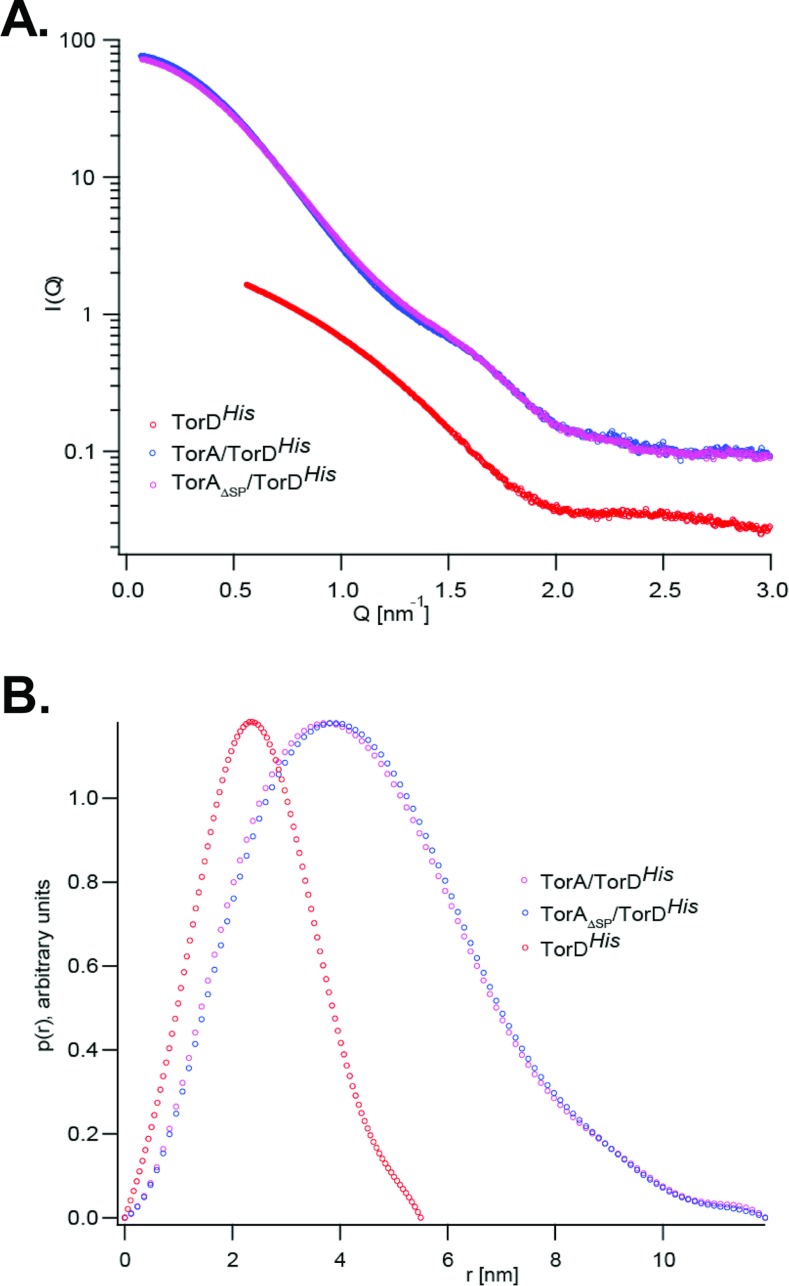
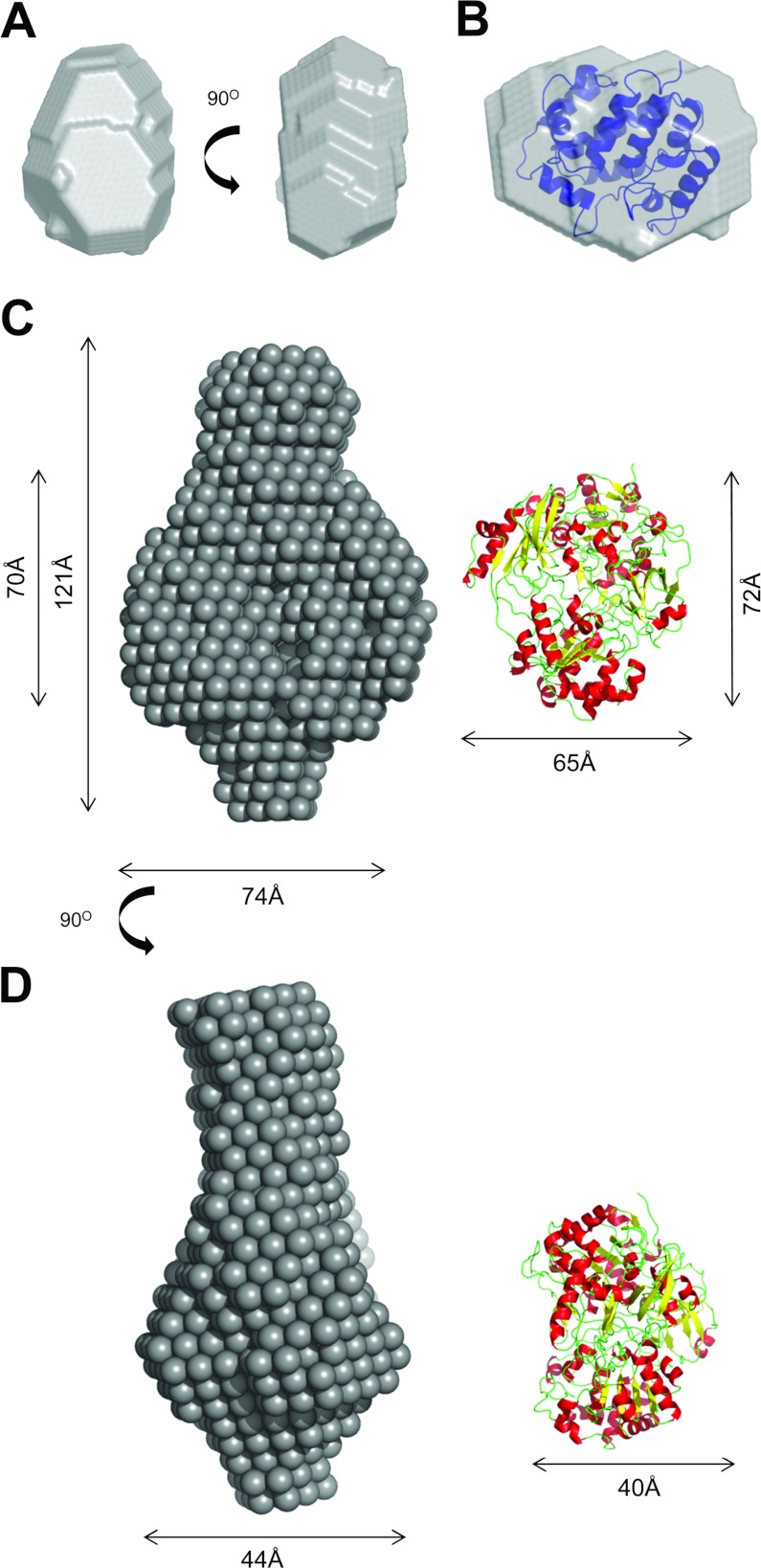
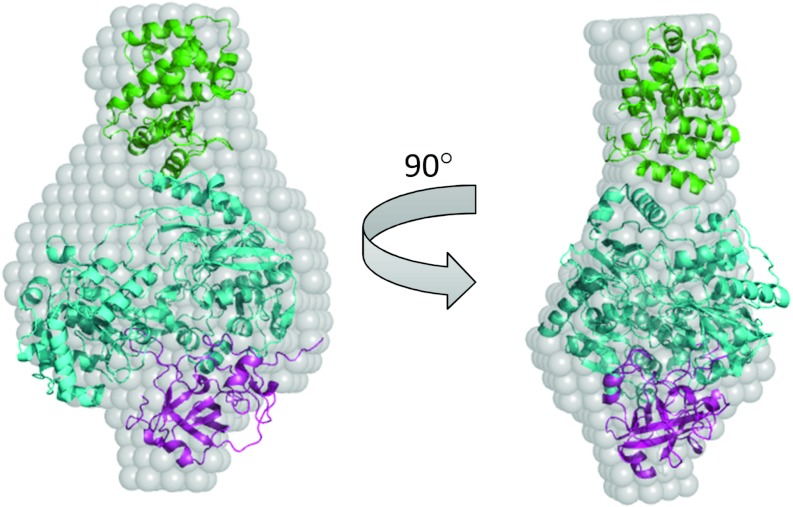
The Tat (twin-arginine translocation) system is a protein targeting pathway utilized by prokaryotes and chloroplasts. Tat substrates are produced with distinctive N-terminal signal peptides and are translocated as fully folded proteins. In Escherichia coli, Tat-dependent proteins often contain redox cofactors that must be loaded before translocation. Trimethylamine N-oxide reductase (TorA) is a model bacterial Tat substrate and is a molybdenum cofactor-dependent enzyme. Co-ordination of cofactor loading and translocation of TorA is directed by the TorD protein, which is a cytoplasmic chaperone known to interact physically with the TorA signal peptide. In the present study, a pre-export TorAD complex has been characterized using biochemical and biophysical techniques, including SAXS (small-angle X-ray scattering). A stable, cofactor-free TorAD complex was isolated, which revealed a 1:1 binding stoichiometry. Surprisingly, a TorAD complex with similar architecture can be isolated in the complete absence of the 39-residue TorA signal peptide. The present study demonstrates that two high-affinity binding sites for TorD are present on TorA, and that a single TorD protein binds both of those simultaneously. Further characterization suggested that the C-terminal 'Domain IV' of TorA remained solvent-exposed in the cofactor-free pre-export TorAD complex. It is possible that correct folding of Domain IV upon cofactor loading is the trigger for TorD release and subsequent export of TorA.
Figures
References
-
- Palmer T., Berks B. C. The twin-arginine translocation (Tat) protein export pathway. Nat. Rev. Microbiol. 2012;10:483–496. - PubMed
-
- Berks B. C. A common export pathway for proteins binding complex redox cofactors? Mol. Microbiol. 1996;22:393–404. - PubMed
-
- Palmer T., Sargent F., Berks B. C. The Tat protein export pathway. In: Böck A., Curtiss R., Kaper J. B., Karp P. D., Neidhardt F. C., Nyström T., Slauch J. M., Squires C. L., Ussery D., editors. EcoSal–Escherichia coli and Salmonella: Cellular and Molecular Biology. Washington, DC: ASM Press; 2010. Chapter 4.3.2, http://www.ecosal.org.
-
- Sargent F. Constructing the wonders of the bacterial world: biosynthesis of complex enzymes. Microbiology. 2007;153:633–651. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases