

Encoding and decoding cellular information through signaling dynamics
- PMID: 23452846
- PMCID: PMC3707615
- DOI: 10.1016/j.cell.2013.02.005
Encoding and decoding cellular information through signaling dynamics
Abstract
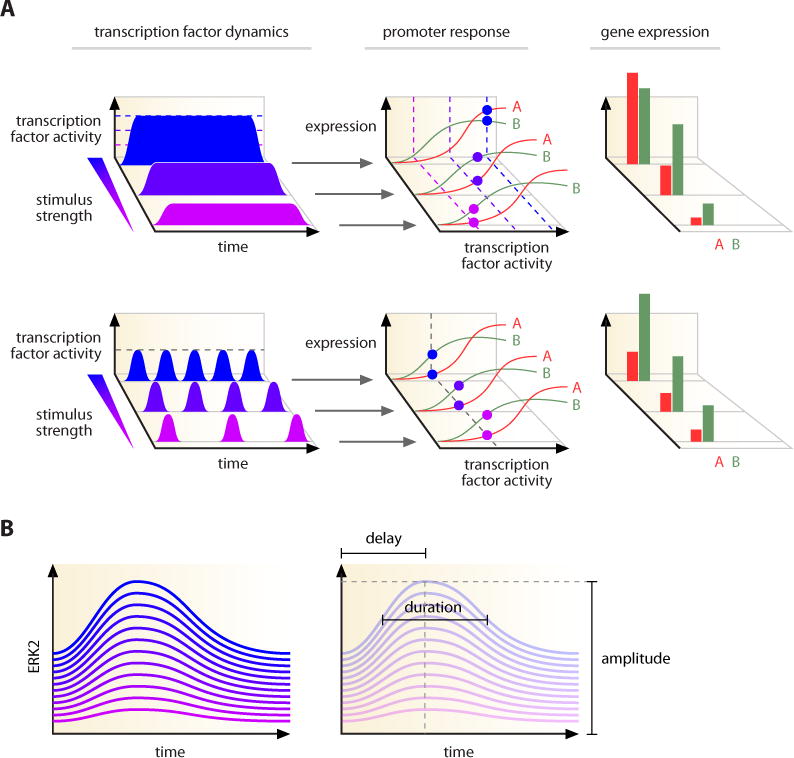
A growing number of studies are revealing that cells can send and receive information by controlling the temporal behavior (dynamics) of their signaling molecules. In this Review, we discuss what is known about the dynamics of various signaling networks and their role in controlling cellular responses. We identify general principles that are emerging in the field, focusing specifically on how the identity and quantity of a stimulus is encoded in temporal patterns, how signaling dynamics influence cellular outcomes, and how specific dynamical patterns are both shaped and interpreted by the structure of molecular networks. We conclude by discussing potential functional roles for transmitting cellular information through the dynamics of signaling molecules and possible applications for the treatment of disease.
Copyright © 2013 Elsevier Inc. All rights reserved.
Figures





References
-
- Alon U. Network motifs: theory and experimental approaches. Nat Rev Genet. 2007;8:450–461. - PubMed
-
- Amit I, Citri A, Shay T, Lu Y, Katz M, Zhang F, Tarcic G, Siwak D, Lahad J, Jacob-Hirsch J, et al. A module of negative feedback regulators defines growth factor signaling. Nat Genet. 2007;39:503–512. - PubMed
-
- Bakstad D, Adamson A, Spiller DG, White MR. Quantitative measurement of single cell dynamics. Curr Opin Biotechnol. 2012;23:103–109. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
