

Synaptic potentiation in the nociceptive amygdala following fear learning in mice
- PMID: 23452928
- PMCID: PMC3606120
- DOI: 10.1186/1756-6606-6-11
Synaptic potentiation in the nociceptive amygdala following fear learning in mice
Abstract
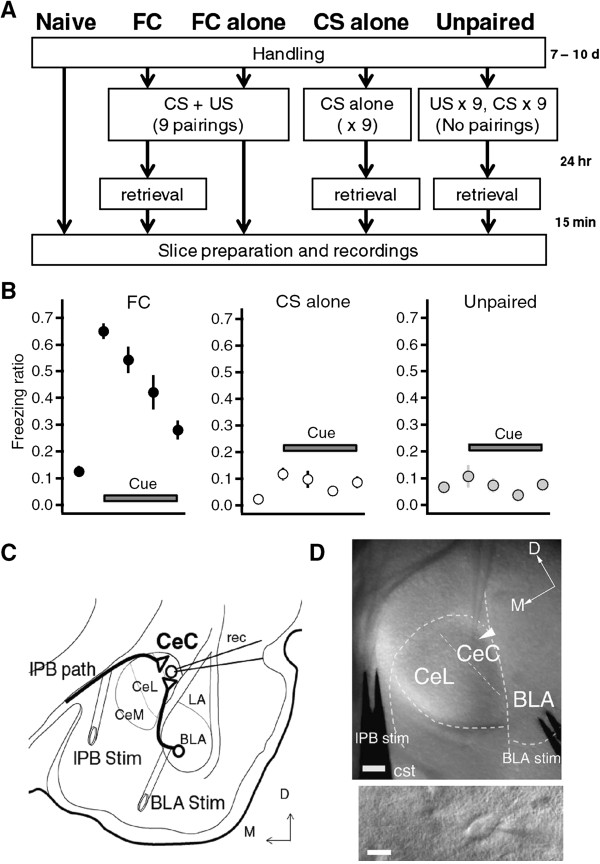
Background: Pavlovian fear conditioning is a classical form of associative learning, which depends on associative synaptic plasticity in the amygdala. Recent findings suggest that the central amygdala (CeA) plays an active role in the acquisition of fear learning. However, little is known about the synaptic properties of the CeA in fear learning. The capsular part of the central amygdala (CeC) receives direct nociceptive information from the external part of the lateral parabrachial nucleus (lPB), as well as highly processed polymodal signals from the basolateral nucleus of the amygdala (BLA). Therefore, we focused on CeC as a convergence point for polymodal BLA signals and nociceptive lPB signals, and explored the synaptic regulation of these pathways in fear conditioning.
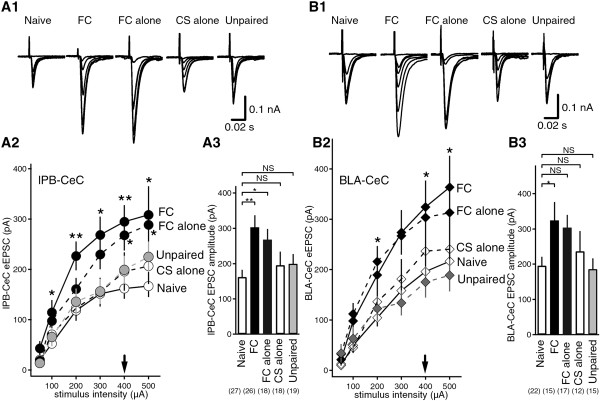
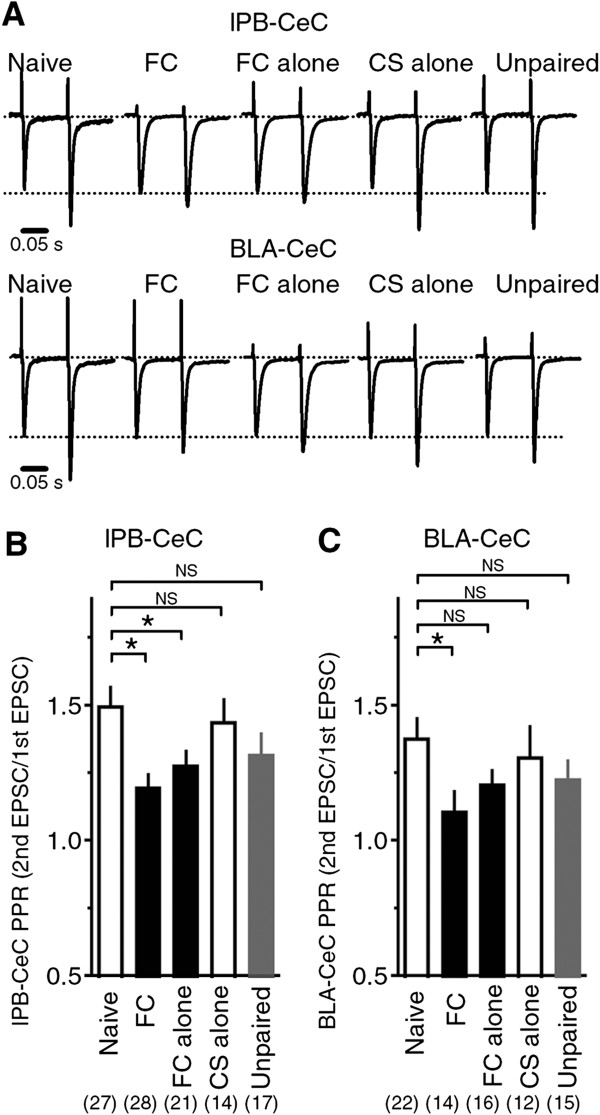
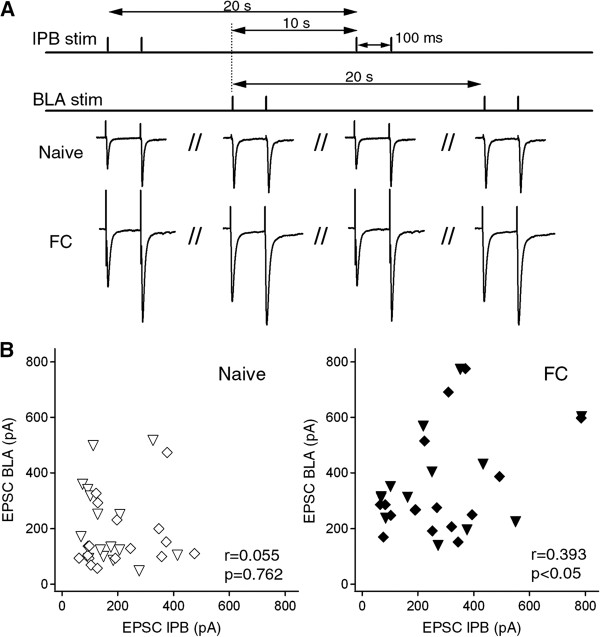
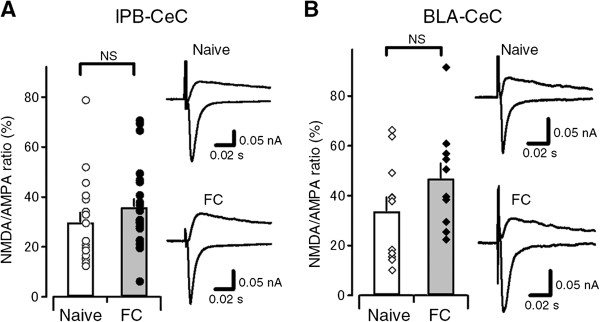
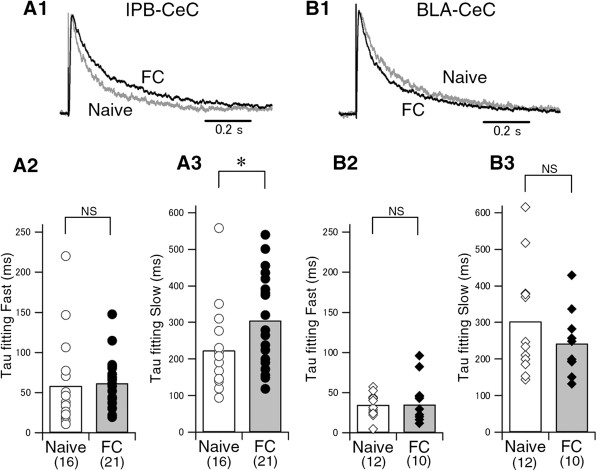
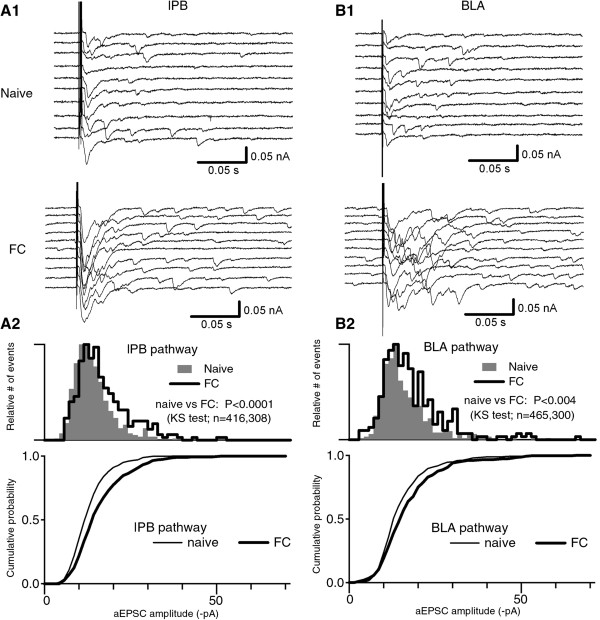
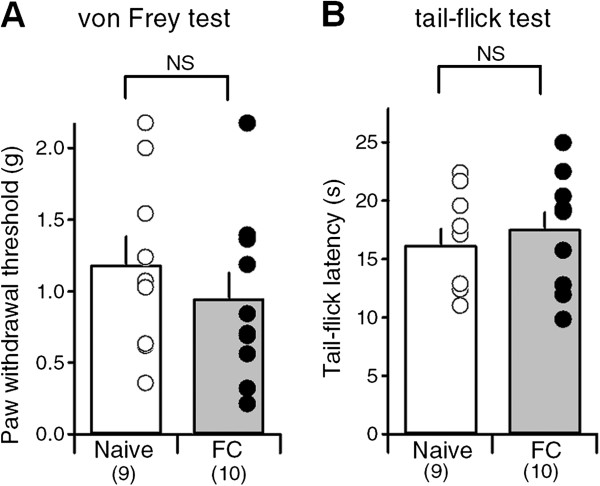
Results: In this study, we show that fear conditioning results in synaptic potentiation in both lPB-CeC and BLA-CeC synapses. This potentiation is dependent on associative fear learning, rather than on nociceptive or sensory experience, or fear memory retrieval. The synaptic weight of the lPB-CeC and BLA-CeC pathways is correlated in fear-conditioned mice, suggesting that fear learning may induce activity-dependent heterosynaptic interactions between lPB-CeC and BLA-CeC pathways. This synaptic potentiation is associated with both postsynaptic and presynaptic changes in the lPB-CeC and BLA-CeC synapses.
Conclusions: These results indicate that the CeC may provide an important locus of Pavlovian association, integrating direct nociceptive signals with polymodal sensory signals. In addition to the well-established plasticity of the lateral amygdala, the multi-step nature of this association system contributes to the highly orchestrated tuning of fear learning.
Figures
References
-
- Wilensky AE, Schafe GE, Kristensen MP, LeDoux JE. Rethinking the fear circuit: the central nucleus of the amygdala is required for the acquisition, consolidation, and expression of Pavlovian fear conditioning. J Neurosci. 2006;26(48):12387–12396. doi: 10.1523/JNEUROSCI.4316-06.2006. - DOI - PMC - PubMed
-
- Busti D, Geracitano R, Whittle N, Dalezios Y, Manko M, Kaufmann W, Satzler K, Singewald N, Capogna M, Ferraguti F. Different fear states engage distinct networks within the intercalated cell clusters of the amygdala. J Neurosci. 2011;31(13):5131–5144. doi: 10.1523/JNEUROSCI.6100-10.2011. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
