

MEK1 is required for PTEN membrane recruitment, AKT regulation, and the maintenance of peripheral tolerance
- PMID: 23453810
- PMCID: PMC3625979
- DOI: 10.1016/j.molcel.2013.01.037
MEK1 is required for PTEN membrane recruitment, AKT regulation, and the maintenance of peripheral tolerance
Abstract
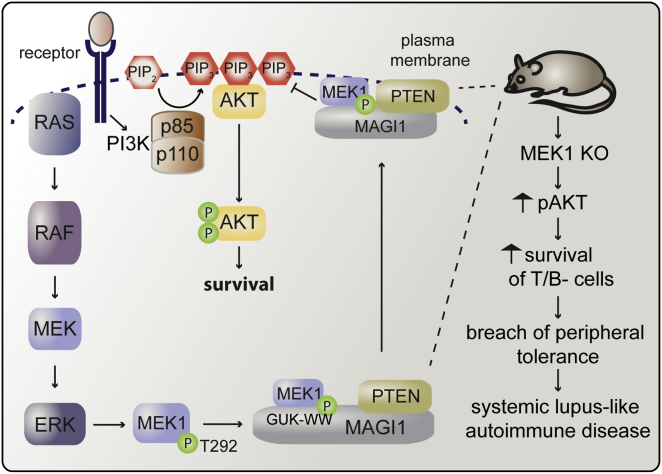
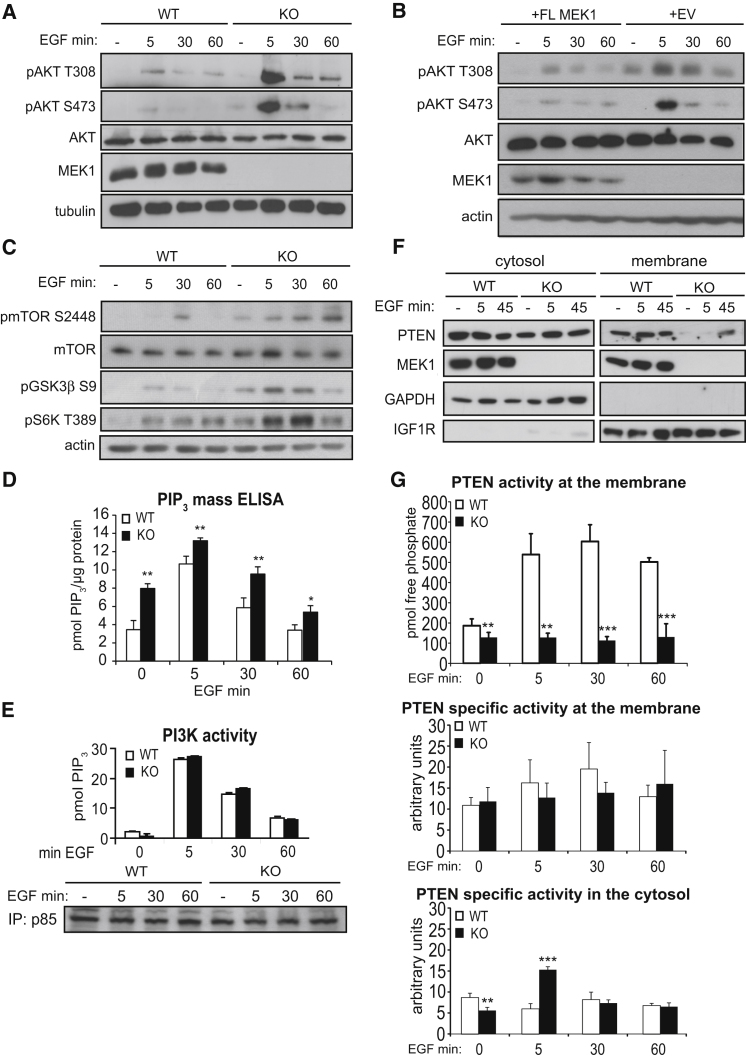
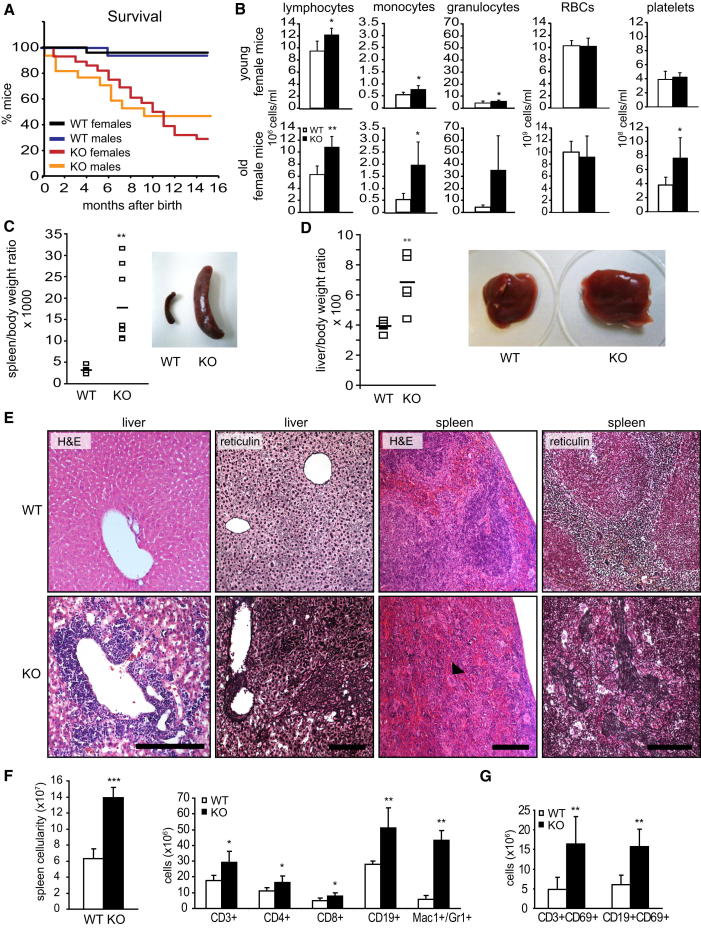
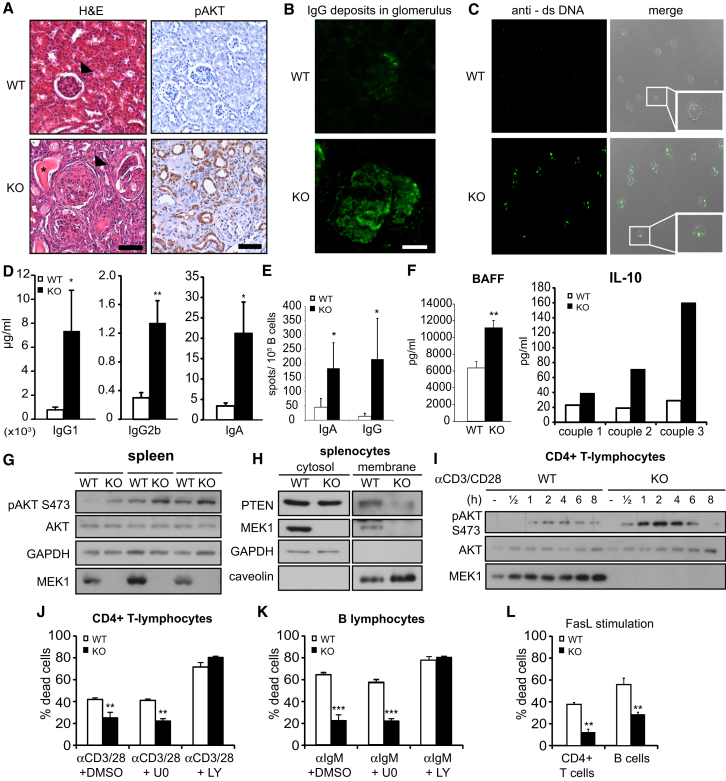
The Raf/MEK/ERK and PI3K/Akt pathways are prominent effectors of oncogenic Ras. These pathways negatively regulate each other, but the mechanism involved is incompletely understood. We now identify MEK1 as an essential regulator of lipid/protein phosphatase PTEN, through which it controls phosphatidylinositol-3-phosphate accumulation and AKT signaling. MEK1 ablation stabilizes AKT activation and, in vivo, causes a lupus-like autoimmune disease and myeloproliferation. Mechanistically, MEK1 is necessary for PTEN membrane recruitment as part of a ternary complex containing the multidomain adaptor MAGI1. Complex formation is independent of MEK1 kinase activity but requires phosphorylation of T292 on MEK1 by activated ERK. Thus, inhibiting the ERK pathway reduces PTEN membrane recruitment, increasing phosphatidylinositol-3-phosphate accumulation and AKT activation. Our data offer a conceptual framework for the observation that activation of the PI3K pathway frequently mediate resistance to MEK inhibitors and for the promising results obtained by combined MEK/PI3K inhibition in preclinical cancer models.
Copyright © 2013 Elsevier Inc. All rights reserved.
Figures
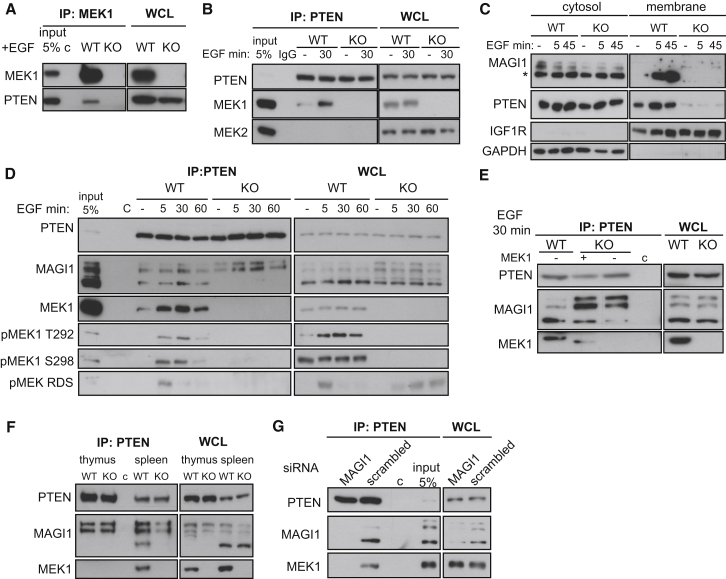
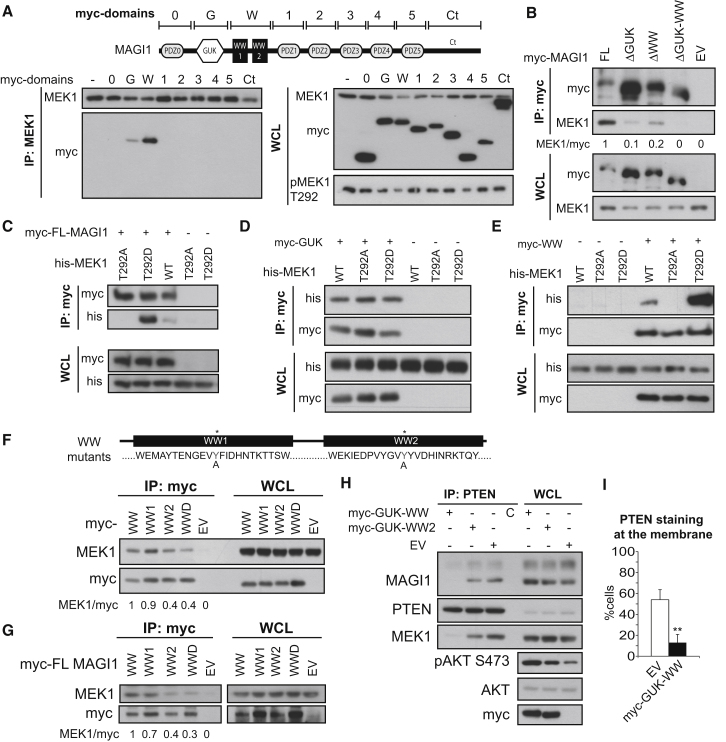
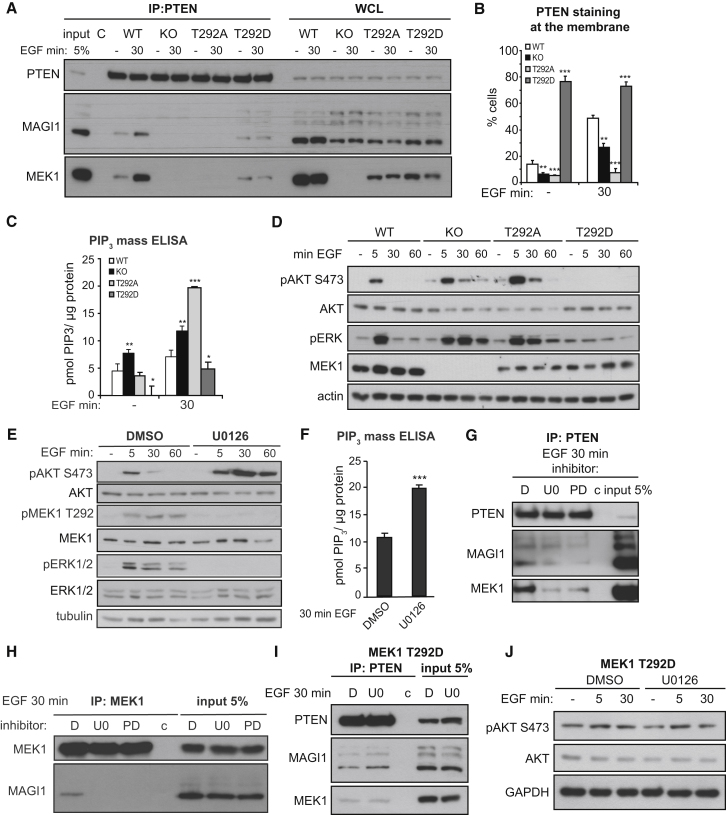
References
-
- Bissonauth V., Roy S., Gravel M., Guillemette S., Charron J. Requirement for Map2k1 (Mek1) in extra-embryonic ectoderm during placentogenesis. Development. 2006;133:3429–3440. - PubMed
-
- Borlado L.R., Redondo C., Alvarez B., Jimenez C., Criado L.M., Flores J., Marcos M.A., Martinez-A C., Balomenos D., Carrera A.C. Increased phosphoinositide 3-kinase activity induces a lymphoproliferative disorder and contributes to tumor generation in vivo. FASEB J. 2000;14:895–903. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
