

Evolution of mir-92a underlies natural morphological variation in Drosophila melanogaster
- PMID: 23453955
- PMCID: PMC3605577
- DOI: 10.1016/j.cub.2013.02.018
Evolution of mir-92a underlies natural morphological variation in Drosophila melanogaster
Abstract
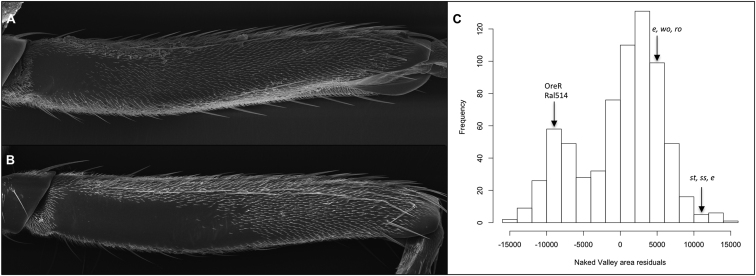
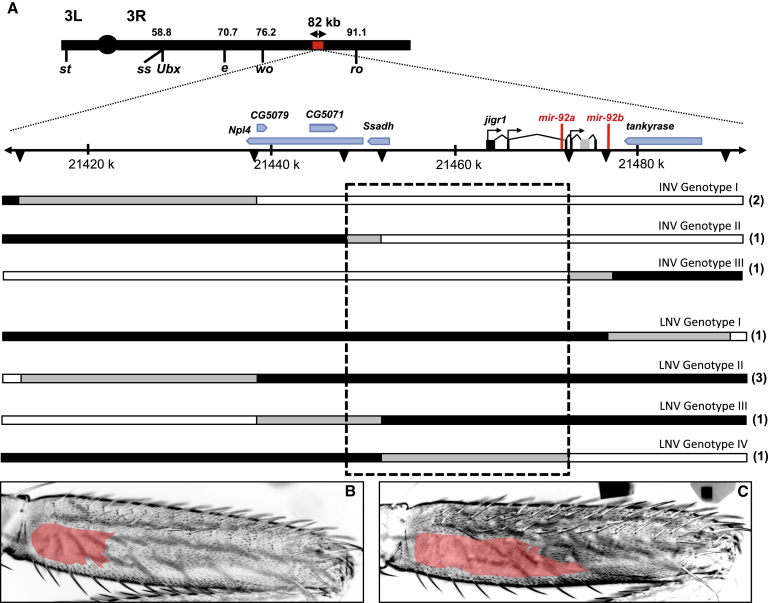
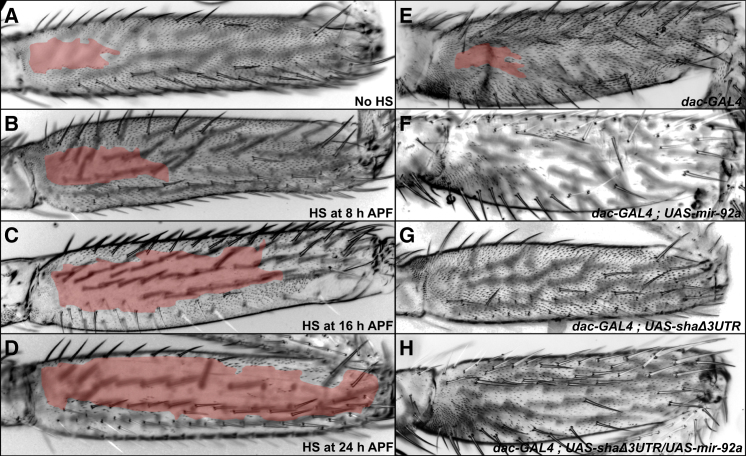
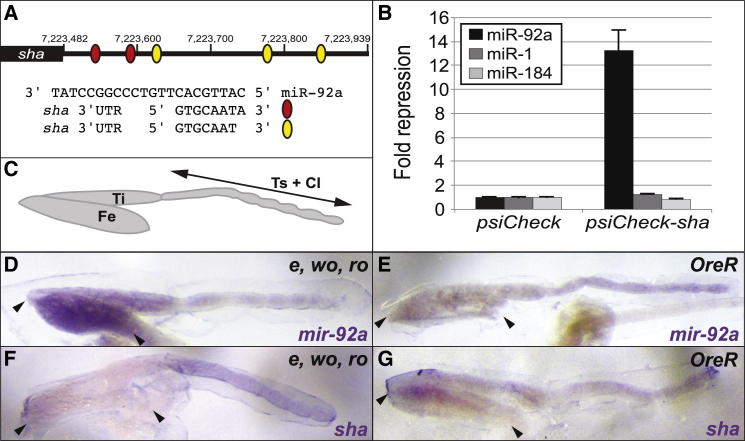
Identifying the genetic mechanisms underlying phenotypic change is essential to understanding how gene regulatory networks and ultimately the genotype-to-phenotype map evolve. It is recognized that microRNAs (miRNAs) have the potential to facilitate evolutionary change [1-3]; however, there are no known examples of natural morphological variation caused by evolutionary changes in miRNA expression. Therefore, the contribution of miRNAs to evolutionary change remains unknown [1, 4]. Drosophila melanogaster subgroup species display a portion of trichome-free cuticle on the femur of the second leg called the "naked valley." It was previously shown that Ultrabithorax (Ubx) is involved in naked valley variation between D. melanogaster and D. simulans [5, 6]. However, naked valley size also varies among populations of D. melanogaster, ranging from 1,000 up to 30,000 μm(2). We investigated the genetic basis of intraspecific differences in the naked valley in D. melanogaster and found that neither Ubx nor shavenbaby (svb) [7, 8] contributes to this morphological difference. Instead, we show that changes in mir-92a expression underlie the evolution of naked valley size in D. melanogaster through repression of shavenoid (sha) [9]. Therefore, our results reveal a novel mechanism for morphological evolution and suggest that modulation of the expression of miRNAs potentially plays a prominent role in generating organismal diversity.
Copyright © 2013 Elsevier Ltd. All rights reserved.
Figures
Comment in
-
Evolutionary genetics: big effect of a small RNA.Curr Biol. 2013 Mar 18;23(6):R247-9. doi: 10.1016/j.cub.2013.02.029. Curr Biol. 2013. PMID: 23518058
References
-
- Li J., Zhang Z. miRNA regulatory variation in human evolution. Trends Genet. 2013;29:116–124. - PubMed
-
- Niwa R., Slack F.J. The evolution of animal microRNA function. Curr. Opin. Genet. Dev. 2007;17:145–150. - PubMed
-
- Alonso C.R., Wilkins A.S. The molecular elements that underlie developmental evolution. Nat. Rev. Genet. 2005;6:709–715. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
